 2015, Vol. 26
2015, Vol. 26扩展功能
文章信息
- 热娜古丽·艾合麦提, 李叶, 张翔, 时磊
- Renagül Exmet, LI Ye, ZHANG Xiang, SHI Lei
- 阿尔金山国家级自然保护区高原鼠兔的食性显微组织分析
- Microscopic histological analysis of diet of Ochotona curzoniae in the Altun Mountain National Nature Reserve
- 中国媒介生物学及控制杂志, 2015, 26(2): 164-167
- Chin J Vector Biol & Control, 2015, 26(2): 164-167
- 10.11853/j.issn.1003.4692.2015.02.014
-
文章历史
- 收稿日期:2014-10-27




2 中国科学院动物研究所;
3 阿尔金山国家级自然保护区管理局
2 Institute of Zoology, Chinese Academy of Sciences;
3 Administrative Bureau of the Altun Mountain National Nature Reserve
食性是哺乳动物种群生物学和生态学研究的核心问题之一[1]。有多种方法可确定植食性哺乳动物的食物[2],其中使用最广泛的间接方法是显微组织鉴别胃内容物或粪便中的植物表皮碎片[3, 4]。啮齿动物食性分析常用的是胃内物,且多为定性分析[5, 6],基于植物显微结构的定量组织分析尚不多见[7]。
高原鼠兔(Ochotona curzoniae)为青藏高原的特有动物,广泛分布于青藏高原的各类草甸。高原鼠兔既是我国青藏高原地区草原主要害鼠之一,也可为其他高原物种提供栖息洞穴和食物资源,因此是高原生态系统的关键种。国内学者对高原鼠兔的食性和食物选择研究涉及食物的主要成分和食量[8, 9]、食物选择模式[10]、不同季节[11]和生境[12]的食性变化,以及达乌尔鼠兔(Ochotona daurica)与高原鼠兔在栖息地的选择、食物资源的竞争等[13, 14]。上述研究多采用直接观察、贮草堆分析或胃内容物显微组织学方法等,尚未见粪便显微组织分析的报道。此外,高原鼠兔作为阿尔金山自然保护区的优势小型哺乳动物[15],广泛分布于各种生境,但主要是高寒荒漠,其栖息地植物群落构成和典型的青藏高原高寒草甸差异明显,对这一地区的草食动物的食性研究尚未见报道。本研究采用粪便显微组织分析法研究阿尔金山自然保护区高原鼠兔不同季节的食物组成,分析主要食物种类的比例及营养生态位,对高原鼠兔的科学管理提供基础资料。
1 材料与方法 1.1 粪样高原鼠兔粪样采集自阿尔金山国家级自然保护区,研究区域自然概况详见文献[15]。将暖季(2013年7月)和冷季(2014年1月)野外采集的各60份粪样按每季10个复合样本进行组合。合计120份20个复合样本,每个复合样本制备5张显微装片。
1.2 样本的制备采集研究地区植物进行种类鉴定,制备46种植物的显微参考装片[16, 17]。高原鼠兔粪样显微装片的制备方法依据文献[18]。
1.3 显微镜片的镜检取粪样装片,每张装片在光学显微镜下检查20个视野(避免重复),放大倍数为100,每个季节合计观察1000个视野。观察每个视野中出现的可辨认植物表皮角质碎片,根据不同的细胞形态类别和特点,参照植物显微装片鉴定出该植物的科或属(种),并记录在视野中出现的次数。
1.4 统计学处理统计求得每种植物角质碎片的出现频率(F),采用频率转换法,依公式:
F=100(1-e-D)
转换为每个视野中每种植物可辨认表皮角质碎片的平均颗粒密度(D),D又可转换为相对颗粒密度RD。方法:
RD=每种植物可辨认的表皮角质碎片的密度/各种植物可辨认角质碎片的密度之和×100%。
RD为食物中各种植物的干重组成比例。即该种植物占食物组成的百分比,据此得出鼠兔食物组成比例。
Shannon-wiener 多样性指数(H):
H=-ΣPiln(Pi)
Pi代表在全部植物碎片中植物物种i的碎片数所占的比例。
Pielou均匀性指数(J): J=H/Hmax
Hmax=lnS,S代表粪样中存在的植物物种数。
食物生态位宽度指数(B): B=1/ΣP2i
用列联表分析高原鼠兔不同季节食物组成比例的差异[19]。所有数据分析在Excel 2003和SPSS 19.0软件中完成。P<0.05为差异有统计学意义。
2 结 果 2.1 食性组成显微组织分析结果显示高原鼠兔暖季采食14科24属25种植物,采食较多(>10%)的植物依次为沙生针茅(Stipa glareosa)、长毛棘豆(Cxytrcpis hisutiuscula)和多花苔草(Carex karoi),3种合计33.58%;按科统计,主要为禾本科(19.15%)、莎草科(15.57%)、豆科(14.29%)和菊科(12.35%)。高原鼠兔冷季共采食12科16属17种植物,采食较多(>10%)的植物依次为沙生针茅、多花苔草、长毛棘豆和垫状驼绒藜(Ceratocarpus compacta),4种合计53.22%;按科统计,主要为禾本科(21.94%)、莎草科(18.53%)、豆科(16.42%)、藜科(10.46%)和菊科(10.32%)(表 1)。按牧草类群统计,相比暖季,高原鼠兔冷季对豆科牧草的采食量增加,而禾草和杂类草比例下降(图 1)。

|
| 图 1 高原鼠兔食物的主要类群 Figure 1 Phylogenetidendrogramof2Bartonellaisolatesconstructe |
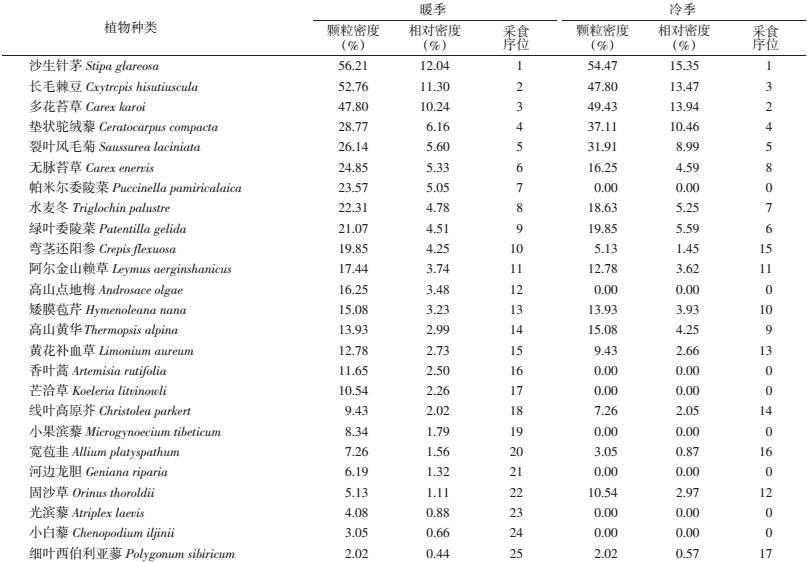
高原鼠兔暖季和冷季采食同种食物的比例不同,且差异有统计学意义(χ2=719.55,df=24,P=0.000,表 2)。不同季节食物采食比例之间存在线性关联。
 |
从表 3可见,高原鼠兔食物种数和生态位宽度暖季大于冷季,而多样性和均匀度暖季小于冷季,表明高原鼠兔冷季采食各种植物比较平均。

啮齿类的食性可以通过多种方法获得,本研究采用粪便显微组织分析法。检出结果实际上是野外条件下食物的丰富程度、鼠兔对食物的喜食程度乃至栖息环境等综合作用的结果,它体现了高原鼠兔在野外条件下实际的食物资源利用状况。食物资源利用分析是了解种群动态的核心,有必要采用非损伤性的方法,以避免对种群动态的干扰。粪便显微组织分析是一种非损伤性的食性分析方法。研究表明胃及结肠内容物组织分析反映了柯氏鼠兔(Ochotona koslowi)在短时间内采食的植物种类,而粪便样品则包含了更多的食物构成信息[20]。尽管食性复杂的物种由于近缘植物显微结构差异微小难以鉴定到种的水平而受到限制[21, 22],但对于食物资源多样性低的区域是一种合适的方法,植物碎片的误定率低且适用于长期监测种群的食性变化[23]。在本研究中,46种参考植物中超过一半(54.35%)出现在粪样中,分属11科18属25种。当地较低的植物多样性及高原鼠兔主要食物植物种间显微结构差异明显,表明粪便显微组织分析对本研究而言是合适的方法。
3.2 高原鼠兔的食性鼠兔(Ochotona spp.)通常被认为是广食性物种[24]。然而,在特定地点,鼠兔优先采食一些植物而忽略了其他广泛可用的植物种类,表现出一定的选择性[25, 26]。国内对高原鼠兔食性的研究表明其食物组成随季节和生境而发生变化。在青藏高原高寒草甸,垂穗披碱草(Elymus nutans)、甘肃棘豆(Oxytropis kansuensis)、铺散亚菊(Ajania khartensis)等是高原鼠兔优先选择的食物[9, 10, 11, 12]。不同栖息地高原鼠兔食物组成的差异主要表现在杂类草草甸,夏季由于喜食种类禾本科和豆科植物资源量减少,高原鼠兔转而大量采食弱小火绒草(Leontopod iumpusillum)、兰石草(Lancea tibetica)和鹅绒萎陵菜(Potentilla anserine)等在矮蒿草草甸和垂穗披碱草草甸不喜食或喜食程度相对较低的杂类草[11];冬季主要采食铺散亚菊和长茎藁本(Ligusticum thomsonii)[12]。在退化草原弃耕地,高原鼠兔主要以紫花针茅(Stipa purpurea)、早熟禾(Poa spp.)、甘肃棘豆等为食[13];在高山蒿草草甸和高山灌丛草甸,高原鼠兔秋季贮存细叶亚菊(Ajania tenuifolia)、甘肃马先蒿(Pedicularis kansuensis)等[27]。扣笼实验发现,随着高原鼠兔种群密度的增大,喜食植物资源量的减少,转而采食平时不太喜食的蒿草属(Kobresia)、苔草属(Carex)和羊茅属(Festuca)植物,说明种内竞争压力的增加,使高原鼠兔的生态位由其偏好资源位构成的核心部分向外围部分泛化[9]。在从垂穗披碱草草甸至矮蒿草草甸和杂类草草甸的生境变化过程中,实质上高原鼠兔喜食食物资源呈现逐渐减少的趋势。尤其杂类草草甸中高原鼠兔喜食的禾本科植物和豆科植物资源较低,导致对食物资源竞争的加剧,食物选择呈现泛化趋势[11]。我们的研究也表明在阿尔金山国家级自然保护区高寒荒漠生境,高原鼠兔主要采食禾本科和豆科牧草,由于植物群落组成差异,其主要采食物种也表现出地理变化。西伯利亚鼠兔(O. hyperborea)[5]和北美鼠兔(O. princeps)[24]亦具有相同的食物选择模式,这一结果也说明栖息地食物可利用量决定了高原鼠兔的食物选择模式。从季节变化看,高原鼠兔暖季的食物生态位宽度明显大于冷季,与暖季的食物资源丰富有关。列联表分析表明,在不同季节高原鼠兔采食同种植物的比例不同,可能与植物资源的可用性有关,表明高原鼠兔能灵活地适应食物可获得性的季节变化[28]。另一方面,冷季的均匀度明显大于暖季,表明冷季食物资源匮乏时,高原鼠兔表现出一定的食性泛化倾向。
植食性哺乳动物的主要食物组成不仅与栖息地的食物可利用量有关,更与食物的质量密切相关,而食物质量则由其营养成分决定。植物的含氮量对草食性动物至关重要,是限制许多草食动物营养摄入的最重要因素。北美鼠兔不根据食物的可获得性贮存植物,而是优先选择那些营养价值高的植物[24];贮草堆中植物的氮含量明显高于生境中最常见的植物[25]。阿尔金山自然保护区不同生境条件下高原鼠兔的食性是否有差异以及和植物营养价值的关系有待进一步研究确认。
志谢 新疆农业大学园林学院王兵副教授帮助鉴定植物标本,动物科学学院研究生王振宇参加部分野外工作,动物科学091班哈斯叶提·艾力参加部分显微实验分析工作,一并志谢.
| [1] | Mátrai K,Altbäcker V,HahnI.Seasonaldietofrabbitsand their browsing effect on juniper in Bugac Juniper Forest (Hungary)[J]. Acta Theriol, 1998, 43:107-112. |
| [2] | 单继红, 吴建平. 食草动物食性研究的主要方法及其评价[J].野生动物杂志, 2005, 26(3):47-49. |
| [3] | 高中信, 陈化鹏, 王筱平. 粪便显微分析法测定植食动物食性的评价[J]. 兽类学报, 1999, 11(3):186-193. |
| [4] | KrisztianK,VilmosA.Diet estimation byfaecesanalysis: sampling optimisation for the European hare[J]. Folia Zool, 2002, 51(1):11-15. |
| [5] | 白雪薇, 史献明, 董国润, 等. 河北省塞北管理区达乌尔黄鼠食性调查[J]. 中国媒介生物学及控制杂志, 2010, 21(4):382-383. |
| [6] | 冯志勇, 邱俊荣, 姚丹丹, 等. 自然条件下板齿鼠的食物结构研究[J]. 中国媒介生物学及控制杂志, 2010, 21(1):23-25. |
| [7] | 雍仲禹, 张美文, 郭聪, 等. 洞庭湖区东方田鼠夏季迁移期在防洪堤坝上的食性[J]. 中国媒介生物学及控制杂志, 2012, 23(1):10-14. |
| [8] | 皮南林. 高原鼠兔的食性及食量研究[M]//灭鼠和鼠类生物学研究报告. 第1集. 北京:科学出版社, 1973:91-102. |
| [9] | 蒋志刚, 夏武平. 高原鼠兔食物资源利用的研究[J]. 兽类学报, 1985, 5(4):251-262. |
| [10] | 王溪, 刘季科, 刘伟, 等. 植食性小哺乳类营养生态学的研究: 高原鼠兔的食物选择模式与食物质量[J]. 兽类学报, 1992, 12(3):183-192. |
| [11] | 刘伟, 张毓, 王溪, 等. 植物生长季节不同栖息地高原鼠兔的食物选择[J]. 兽类学报, 2008, 28(4):358-366. |
| [12] | 刘伟, 张毓, 王溪, 等. 高原鼠兔冬季的食物选择[J]. 兽类学报, 2009, 29(1):12-19. |
| [13] | 樊乃昌, 景增春, 张道川. 高原鼠兔与达乌尔鼠兔食物资源维生态位的研究[J]. 兽类学报, 1995, 15(1):36-40. |
| [14] | 樊乃昌, 张道川. 高原鼠兔与达乌尔鼠兔的摄食行为及对栖息地适应性的研究[J]. 兽类学报, 1996, 16(1):48-53. |
| [15] | 李叶, 王振宇, 张翔, 等. 阿尔金山自然保护区高原鼠兔夏季微生境选择的主导因子分析[J]. 中国媒介生物学及控制杂志, 2014, 25(1):28-31. |
| [16] | 刘伟, 杨维康,徐文轩. 蒙古野驴的秋季食性分析[J]. 兽类学报, 2008, 28(1):33-36. |
| [17] | 李叶, 王兵, 黄永丽, 等. 天山地区三种自由放牧家畜食性和营养生态位的显微组织分析[J]. 家畜生态学报, 2013, 34(9):65-68. |
| [18] | 曹伊凡, 苏建平. 一种用于食草动物粪便显微组织分析的临时装片新技术[J]. 兽类学报, 2006, 26(4):407- 410. |
| [19] | 崔庆虎, 连新明, 张同作, 等. 青海门源地区大鵟和雕鸮的食性比较[J]. 动物学杂志, 2003, 38(6):57-63. |
| [20] | 曹伊凡, 林恭华, 卢学峰, 等. 柯氏鼠兔的食性分析[J]. 动物学杂志, 2009, 44(1):58-62. |
| [21] | Soininen EM,Valentini A,Coissac E,etal.Analysingdiet of small herbivores: the efficiency of DNA barcoding coupled with high-throughput pyrosequencing for deciphering the composition of complex plant mixtures[J]. Front Zool, 2009, 6:16. |
| [22] | Soininen EM, Ravolainen VT,Bråthen KA,et al. Arctic Small rodents have Diverse diets and flexible food selection[J]. PLoS One, 2013, 8(6):e68128. |
| [23] | Chapuis JL, Boussés P,Pisanu B,et al. Comparative rumen and fecal diet microhistological determinations of European mouflon [J]. J Range Manage, 2001, 54(3):239-242. |
| [24] | SmithAT, WestonML. Ochotonaprinceps[J]. Mammalian Species, 1990, 352:1-8. |
| [25] | Gliwicz J, Pagacz S, Witczuk J. Strategy of food plant selection in the siberian Northern Pika, Ochotona hyperborea[J]. Arct Antarct Alp Res, 2006, 38(1):54-59. |
| [26] | Smith JA,Erb LP. Patternsofselectivecachingbehaviorof a generalist herbivore,the American pika (Ochotona princeps) [J]. Arct Antarct Alp Res, 2013, 45(3):396-403. |
| [27] | 张毓, 刘伟, 王学英. 高原鼠兔贮草行为初探[J]. 动物学研究, 2005, 26(5):479-483. |
| [28] | Freschi P,Fascetti S,Musto M,et al. Diet of the apennine hare in a southern Italy Regional Park[J]. Eur J Wildlife Res, 2014, 60(3):423-430. |