 2015, Vol. 26
2015, Vol. 26扩展功能
文章信息
- 廖力夫, 乌守巴特, 章建民, 燕顺生
- LIAO Li-fu, BATE Wu-so, ZHANG Jian-min, YAN Shun-sheng
- 塔里木盆地荒漠型黑热病传播媒介吴氏白蛉的供血宿主调查及分析
- Investigation and analysis on the blood feeding hosts for vector Phlebotomus wui in desert-type kala-azar of Tarim basin
- 中国媒介生物学及控制杂志, 2015, 26(2): 151-154
- Chin J Vector Biol & Control, 2015, 26(2): 151-154
- 10.11853/j.issn.1003.4692.2015.02.011
-
文章历史
- 收稿日期:2014-10-28




2 新疆维吾尔自治区疾病预防控制中心;
3 新疆巴楚县50团疾病预防控制中心
2 Xinjiang Center for Disease Control and Prevention;
3 Center for Disease Control and Prevention of 50 Tuan
媒介型自然疫源性疾病疫源地的维持必须具备3个条件,即病原体、传播媒介和储存宿主[1]。在塔里木盆地荒漠型黑热病自然疫源地中,已查明病原体是利什曼原虫[2, 3],传播媒介是吴氏白蛉(Phlebotomus wui)[4, 5],储存宿主(即传染源)虽然经多年调查,目前只从塔里木兔(Lepus yarkandensis)体内分离出利什曼原虫[6],但由于进行传播媒介感染塔里木兔的实验难度大,而传播媒介感染储存宿主又必须通过吸血来完成,因此调查传播媒介吴氏白蛉的供血宿主是间接证明传播媒介感染储存宿主的有效手段。
为查明吴氏白蛉的供血动物,本研究以塔里木盆地荒漠型黑热病自然疫源地的传播媒介吴氏白蛉为吸血昆虫,以当地优势野生动物种为吸血对象[7],通过媒介叮咬吸血实验,观察吸血率,查找供血动物,通过传播媒介与供血动物对其栖息环境的相互关系分析,寻找荒漠型黑热病储存宿主的科学依据。
1 材料与方法 1.1 野生动物种类及密度调查①夹日法: 捕鼠工具为小号铁板夹,诱饵为油炸花生米,每天傍晚外生境每5 m间距布放1夹,次日清晨收夹,鉴定动物种类,计算动物密度。②足链法:清晨计数调查区域沙土表面前1天夜晚动物活动留下的新鲜足链,用GPS(全球定位系统)定位测定调查的距离,鉴别不同动物的足迹,计数单位距离的动物足链数。根据调查结果,确定不同生境的优势动物种类。
1.2 受试动物根据白蛉季节查明的优势野生动物种类,夏季于巴楚县夏河林场附近的沙丘用捕鼠笼或鸟网诱捕优势野生动物,或从当地农民处收购,获得动物单笼室内饲养1周以上备用。
1.3 吴氏白蛉在既往调查吴氏白蛉比例达95%以上的新疆巴楚县夏河林场[4],于2005年6-8月白蛉活动高峰期,用灯光引诱,吸蚊器吸捕白蛉。将装有白蛉的吸蚊头移入冰箱冷冻室30 s左右,白蛉低温麻醉后,倒入底部附有冰袋的培养皿内,在解剖镜下将未吸血的雌性吴氏白蛉取出,移入大纸箱内的白蛉饲养笼内备用,饲养环境温度25~27 ℃,相对湿度50%~65%。
1.4 吸血时动物的活动类型动物活动类型分自由活动组和限制活动组。自由活动组:笼内动物可自由活动和取食。体重<100 g的野生鼠类饲养在11 cm×12 cm×22 cm铁丝笼中,笼的网眼? 0.8 cm;体重>200 g的动物饲养在16 cm×18 cm×25 cm铁丝笼中,笼的网眼 2.0 cm。每笼1只。限制活动组:吸血前将野生鼠类移入几乎不能活动的小铁丝笼内,再移入白蛉饲养笼进行实验。
1.5 叮咬吸血实验体重>200 g的动物在20 cm×40 cm×50 cm昆虫笼中进行叮咬吸血实验,体重<100 g的动物在15 cm×20 cm×30 cm昆虫笼中进行叮咬吸血实验。先将装有动物的铁丝笼下垫托盘移入昆虫笼内,封闭昆虫笼,再将饥饿24 h以上的备用白蛉移入昆虫笼内,每次实验用1只动物。叮咬实验于天黑前开始,第2天清晨结束后用吸蚊器吸出笼内所有白蛉,按1.3方法低温麻醉白蛉后,在解剖镜下检查,白蛉肠道有红细胞为吸过血的白蛉,计算白蛉对受试动物的吸血率。叮咬吸血期间,同时观察记录白蛉叮咬的动物部位、吸血时间、动物干扰白蛉叮咬的行为等。
2 结 果 2.1 红柳胡杨生境的优势动物调查结果显示,塔里木盆地荒漠型黑热病自然疫源地不同生境的各种野生动物种类和密度存在明显差异(表 1、表 2),在居民区及其周边,优势野生动物主要是小家鼠(Mus muscullus)、子午沙鼠(Meriones meridianus),在沙包+红柳+胡杨生境,白蛉活动期的优势野生动物主要有子午沙鼠、毛脚跳鼠(Dipus sagitta)、科氏跳鼠(Salpingotus kozlovi)、塔里木兔和大耳猬(Hemiechinus auritus)。
 |
 |
根据红柳胡杨生境调查出的优势动物和白蛉(表 3),对5种动物进行叮咬实验(表 4)。在5种受试的自由活动动物中,吴氏白蛉只能吸到塔里木兔和大耳猬血,吸血率分别为9.5%和3.3%,其他3种啮齿动物的吸血率为0。
 |
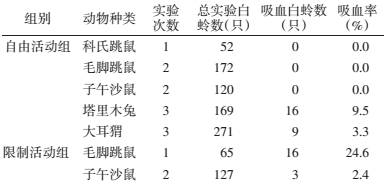
将自由活动吸不到血的动物毛脚跳鼠和子午沙鼠用小铁丝笼限制其肢体活动,结果2种啮齿动物均可以被吴氏白蛉吸到血,吸血率分别为24.6%和2.4%(表 4)。
2.4 吸血部位在自由活动组,吴氏白蛉对塔里木兔的吸血部位在耳内无毛部位,对大耳猬的吸血部位在鼻尖无毛部位,对其他3种啮齿动物,因肢体驱赶活动干扰,白蛉无法接触吸不上血。在限制活动组,因动物的肢体驱赶干扰活动被限制,主要吸血部位集中在2种动物的嘴部毛少部位、耳壳内侧无毛部位和尾部毛少部位。
2.5 吸血时间白蛉移入昆虫笼10 min左右开始吸血。如果无动物肢体驱赶干扰行为,白蛉10~30 min可以吸饱血。如果动物有驱赶活动,白蛉只能不断变换吸血部位,延长吸血时间,吸很少的血,甚至吸不到血。
3 讨 论 3.1 传播媒介供血宿主的判断依据作为黑热病的动物储存宿主,首先能够自然感染利什曼原虫,但前提是动物在自由活动条件下能被传播媒介叮咬吸血,动物才有感染的机会。如果动物能发病并保存,黑热病病原体才能在疫源地持续维持下去。如果传播媒介与储存宿主这一最基本的供血关系都无法保证,也就无法充当利什曼原虫储存宿主的角色[8, 9]。因此动物在自由活动条件下能否被传播媒介叮咬吸血可以作为验证动物是否具备供血动物的重要条件。在塔里木盆地荒漠型黑热病的居民区,人、家畜和野生动物是吴氏白蛉的主要供血宿主[4],在原始荒漠区或荒郊野外,除临时侵入的人或家畜外,当地优势野生动物应该是白蛉的主要吸血选择对象(供血宿主)。供血宿主的选择取决于吴氏白蛉对每种动物的嗜血性和每种动物的自身特征及习性,如吴氏白蛉对羊和驴的内耳、肛门附近无毛部位吸血[4, 5],以及本实验对塔里木兔内耳的吸血,均与暴露的无毛面积和动物的活动状态有关。
根据本实验对自由活动的5种动物实验,至少可以提供如下证明:①自由活动吸血实验可以找出吴氏白蛉的供血宿主;②吴氏白蛉对实验的受试哺乳动物存在嗜血性;③动物肢体驱赶活动是干扰白蛉嗜吸自由活动动物血的重要原因之一;④限制动物活动的方法可以增加吴氏白蛉对动物的吸血机会。
3.2 黑热病储存宿主的潜伏期和被感染季节文献资料显示,被利什曼原虫感染的白蛉叮咬黑线仓鼠(Cricetulus barabensis)5~7个月后,才有明显的利什曼病临床特征[10],即白蛉叮咬5~7个月后的黑线仓鼠才有传播利什曼原虫的可能性。根据传播媒介吴氏白蛉6-9月的活动季节推测[4, 5],当年被白蛉叮咬感染的动物来年白蛉活动季节(6-9月)才有再感染其他动物的可能性,当年具有传播性的动物基本上是上一年感染利什曼原虫的动物,如果该假设成立,储存宿主的寿命应该>9个月,黑热病自然疫源地才能长期维持下去。因此,储存宿主的寿命越长,自然疫源地维持的基础越牢固。
供血宿主是作为储存宿主的必要条件之一,当供血宿主与传播媒介维持充分的接触机会,被吸血并能感染利什曼原虫维持9个月以上的感染状态,来年传播媒介活动时,供血宿主才有作为储存宿主的可能。
本实验使用的5种动物,3种鼠类的野外平均寿命一般在1年左右[11],塔里木兔和大耳猬的平均寿命>2年[12, 13]。从自由活动状况下5种动物的吸血结果看,吴氏白蛉只吸到塔里木兔和大耳猬血,表明塔里木兔和大耳猬均有感染利什曼原虫的机会,根据前者吸血率是后者吸血率3倍的结果推测(表 4),前者被感染的机会应该大于后者。现场调查的动物抗利什曼原虫抗体阳性率结果也能证实,只有塔里木兔查出抗体阳性(44/177),而大耳猬和其他3种自由活动时白蛉吸不到血的动物抗体均为阴性[6]。
3.3 黑热病患者或储存宿主被感染的环境根据塔里木盆地荒漠型黑热病疫区每年新发现的黑热病患者均有白蛉活动期进入红柳胡杨生境或居住在红柳胡杨生境附近的流行病学经历,以及该生境传播媒介吴氏白蛉的利什曼原虫感染率高于其他环境的结果推测[5, 14, 15],黑热病患者或储存宿主在该生境被感染的可能性最大,该生境栖息的优势动物被吴氏白蛉吸血的可能性也最大。
3.4 传播媒介和供血宿主对栖息环境的选择及其相互关系环境地下水位的高低和微环境温湿度生境是传播媒介吴氏白蛉选择栖息繁殖生境的重要条件[16]。在塔里木盆地荒漠型黑热病疫区,除农田外,荒漠干旱区相对湿度最高的稠密柽柳胡杨生境,是传播媒介吴氏白蛉的理想繁殖和栖息场所[4](表 3)。为防止天敌等捕捉,塔里木兔常选择柽柳胡杨生境栖息藏身和繁殖(表 2),其窝巢常选择于稠密的灌丛中,其幼体生下时就全身被毛双眼睁开,能自由活动取食。塔里木兔的成体和幼体与吴氏白蛉共同栖息的柽柳胡杨生境,为传播媒介与供血宿主创造了共生条件,当吴氏白蛉反复吸食塔里木兔血时,将利什曼原虫从塔里木兔成体转染给幼体,如此代代转染,使荒漠型黑热病自然疫源地的长期维持成为可能。
| [1] | 唐家琪.自然疫源性疾病[M]. 北京:科学出版社, 2005: 237-241. |
| [2] | 郑学礼, 胡孝素, 陈建平. 我国不同流行区内脏利什曼原虫分 离株kDNA的PCR-SSCP分析[J]. 中国寄生虫学与寄生虫病 杂志, 1999, 17(6):345-349. |
| [3] | Matlashewski G,Arana B,Kroeger A,et al. Visceral leishmaniasis: Elimination with existing interventions[J]. Lancet Infect Dis,2011, 11(11):322-325. |
| [4] | 熊光华, 管立人, 王捷, 等. 新疆荒漠硕大白蛉吴氏亚种的生 态习性及其防制[J]. 昆虫学报, 1979, 22(4):428-436. |
| [5] | 熊光华, 管立人, 郭印宽. 新疆黑热病传播媒介的研究[J]. 流 行病防治研究, 1974, 2(4):327-334. |
| [6] | 廖力夫, 燕顺生, 乌守巴特, 等. 从塔里木兔体内首次分离出婴儿利什曼原虫[J]. 中国媒介生物学及控制杂志, 2009, 20 (1):45-47. |
| [7] | 乌守巴特, 燕顺生, 雷刚, 等. 新疆巴楚县夏河林场小型鼠类 及其体外寄生物调查[J]. 地方病通报, 2010, 25(3):5-7. |
| [8] | World Health Organization. Control of the Leishmaniases[M]. Geneva:WHO, 1990:38-40. |
| [9] | Wang JY, Cui G, Chen HT, et al. Current epidemiological profile and features of visceral leishmaniasis in People's Republic of China[J]. Parasit Vectors, 2012, 5:31. |
| [10] | 管立人, 柴君杰, 左新平. 新疆白蛉生物学研究进展[J]. 地方 病通报, 1999, 14(4):87-91. |
| [11] | 王思博, 杨赣源. 新疆啮齿动物志[M]. 乌鲁木齐:新疆人民出 版社, 1983:93, 108-110, 150-154. |
| [12] | 罗泽洵. 中国野兔[M]. 北京:中国林业出版社, 1988:87-103. |
| [13] | Wilson DE, Reeder DM. Mammal species of the world[M]. 3rd. Baltimore MD. Johns Hopkins University Press, 2005:215. |
| [14] | Wang JY, Gao CH, Yang YT, et al. An outbreak of the desert sub-type of zoonotic visceral Leishmaniasis in Jiashi, Xinjiang Uygur Autonomous Region, People's Republic of China[J]. Parasitol Int, 2010, 59(3):331-337. |
| [15] | Narayan RB, Gert VA, Suman R, et al. Domestic animals and epidemiology of visceral Leishmaniasis, Nepal[J]. Emerg Infect Dis, 2010, 16(2):231-237. |
| [16] | 管立人, 柴君杰, 杨利普. 新疆维吾尔自治区白蛉的地域分布 与地理景观的关系[J]. 中国寄生虫学与寄生虫病杂志,1996, 14(1):26-32. |