 2014, Vol. 25
2014, Vol. 25扩展功能
文章信息
- 李海龙, 袁江玲, 张桂林, 郑重, 刘晓明, 孙响
- LI Hai-long, YUAN Jiang-ling, ZHANG Gui-lin, ZHENG Zhong, LIU Xiao-ming, SUN Xiang
- 新疆北部干旱区5个湿地蚊类的生态位及无倾向对应排序分析
- Niche breadth, overlap, and detrended correspondence analysis of mosquitoes in five arid wetlands in North Xinjiang, China
- 中国媒介生物学及控制杂志, 2014, 25(6): 492-495
- Chin J Vector Biol & Control, 2014, 25(6): 492-495
- 10.11853/j.issn.1003.4692.2014.06.002
-
文章历史
- 收稿日期:2014-6-4




2 新疆维吾尔自治区疾病预防控制中心
2 Xinjiang Center for Disease Control and Prevention
对于某一生物种群而言,生态位(niche)的相关指标能够反映出其地理分布情况以及生境的适应性。通常用生态位宽度(niche breadth)、生态位重叠指数(niche overlap)和排序分析来定量地描述生物群落在不同生境的生态位特征以及种群与生境之间的相互关系。新疆维吾尔自治区(新疆)北部干旱区湿地蚊虫危害十分严重,其刺叮骚扰及携带的传染性病原对周边居民的生活和健康造成了较大影响。国内外已经开展较多的关于蚊虫区系分布、生物学习性、生物多样性等一系列研究,但针对新疆地区尤其是干旱区湿地的研究多集中于蚊虫种群组成、活动节律调查、防制措施以及蚊虫的医学重要性等方面[1, 2, 3, 4, 5, 6, 7],而少有涉及关于蚊虫生态位方面的研究[8]。在蚊类生态学方面,国内外学者常利用大尺度生态学、景观生态学进行蚊类群落结构、环境因素对蚊虫种群分布的影响等进行研究[9, 10, 11, 12],通过聚类分析法和排序分析法[13, 14, 15, 16, 17]来研究探讨蚊虫种群间相关性及种群与环境因子之间的内在联系。因此本研究有助于进一步了解干旱区湿地蚊虫群落特点及其与环境之间的相互关系,为干旱区蚊类及蚊媒病防制提供理论依据。 1 材料与方法 1.1 研究区域
选择新疆北部伊犁河、额敏河、乌伦古河分支布尔根河、额尔齐斯河4个河流型湿地以及艾比湖平原湖泊型湿地进行调查。调查样地分别位于伊犁河下游三道河地区(43°50′33″N,80°38′44″E,海拔540 m)、布尔根河塔克什肯地区(46°10′51″N,90°48′47″E,海拔1114 m)、额敏河河口地区(46°28′43″N,82°53′46″E,海拔407 m)、额尔齐斯河下游湿地北湾地区(48°01′51″N,85°32′30″E,海拔417 m)以及艾比湖西侧与农五师九十团毗邻地区(44°48′54″N,82°53′16″E,海拔201 m)。 1.2 研究方法 1.2.1 样地调查
2006-2010年,于6-8月蚊虫活动高峰时间,在每个调查样地,选择居住区内树林、河岸林、盐渍化荒地3种不同典型生境作为标本采集点,采用CO2灯诱及人帐诱法收集蚊虫标本,对标本计数和分类鉴定。 1.2.2 数据处理
生态位宽度(Bi)采用Shannon?Wiener多样性指数为基础的生态位宽度指数[18]。生态位重叠指数(NO)采用Pianka(1973)的重叠指数[18]。应用各湿地生境内不同蚊种的构成比构建矩阵,采用IBM SPSS Statistics 19.0软件进行系统聚类分析。以不同湿地生境内各种蚊虫数量为指标,变量经过Y=lg(y+1)标准化,应用无倾向对应分析(detrended correspondence analysis,DCA)探讨不同生境对蚊虫分布的影响,DCA分析通过Canoco for Windows 4.5软件完成。 2 结 果
通过调查,在5个干旱区湿地共捕获蚊类5属16种共20 731只。其中,伊犁河下游5属8种共5431只;艾比湖平原湖泊型湿地5属11种共4716只;乌伦古河分支布尔根河3属5种共2664只;额敏河下游4属9种共1218只;额尔齐斯河下游4属8种共6702只(表 1)。在所获16种蚊虫中,刺扰伊蚊(Aedes vexans)为优势蚊种,占捕获总数的55.98%,常见种类有里海伊蚊(Ae. caspius)、赫坎按蚊(Anopheles hyrcanus)、凶小库蚊(Culex modestus)、尖音库蚊(Cx. pipiens)和米赛按蚊(An. messeae),分别占13.68%、11.98%、9.03%、4.89%和1.88%,其他蚊种数量较少。
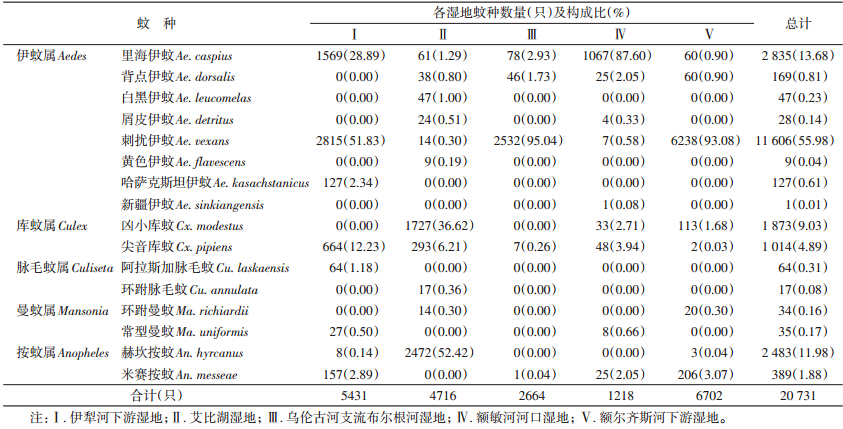 |
根据生态位宽度测定,最大值为背点伊蚊(0.8326),其次为刺扰伊蚊(0.6350)、里海伊蚊(0.5954)和米赛按蚊(0.5558)。生态位重叠指数结果(表 2)显示,NO>0.9000以上的蚊种组分别有刺扰伊蚊与米赛按蚊等;26个种对数NO>0.90,占20.31%,85个种对数NO<0.50,占70.83%。
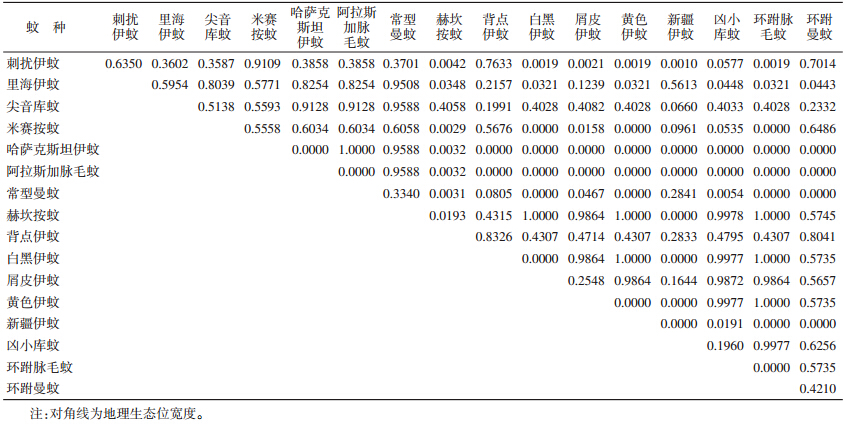 |
通过对5个湿地蚊类构成的系统聚类分析,结果将5个湿地的蚊类生境分为2类。其中,乌伦古河支流布尔根河湿地、额尔齐斯河下游湿地及伊犁河下游3个湿地被聚为一类,而艾比湖湿地和额敏河河口湿地被聚为另一类(图 1)。

|
| 图 1 新疆北疆干旱区5个湿地蚊类系统聚类 Figure 1 Dendrogram of cluster analysis based on the constituent ratio of individual mosquito species in five arid wetlands in North Xinjiang,China |
DCA排序结果表明,新疆伊蚊和环跗曼蚊距离其他蚊种最远。阿拉斯加脉毛蚊与哈萨克斯坦伊蚊相关性最大,分布完全一致;白黑伊蚊、黄色伊蚊、环跗脉毛蚊相关性最大,分布完全一致;刺扰伊蚊与米赛按蚊和里海伊蚊相关性较强;背点伊蚊、凶小库蚊、白黑伊蚊、黄色伊蚊、环跗脉毛蚊呈近似线性,也表现出很强的相关性(图 2)。

|
| 图 2 新疆北疆干旱区5个湿地蚊类DCA二维排序结果 Figure 2 Two?dimensional ordination diagram of DCA for mosquitoes in five arid wetlands in North Xinjiang,China |
生态位宽度指数越大,说明该物种对于环境适应性的特异性越小,分布也就越广泛。但在本研究中作为优势种的刺扰伊蚊,其生态位宽度(0.6350)却低于背点伊蚊(0.8326),其原因可能是刺扰伊蚊在生境中较为集中分布,均匀度较低所致,说明生态位宽度最大的种不一定是优势种。物种在可利用资源丰富的情况下,能选择性地利用资源,使得种群数量分布不均衡,从而使物种生态位宽度变窄[19]。本研究中生态位宽度位于前4位的有刺扰伊蚊、里海伊蚊、米赛按蚊和背点伊蚊,说明这4个蚊种在干旱环境中具有较强的适应性,因此在新疆北部地区的分布范围较广,成为当地的优势种或常见种。
生态位重叠作为衡量种间生态相似性的指标。生态位重叠值高反映的是两种蚊种对生态因子有着相似要求,反之则说明差异性较大,因此,生态位重叠也导致了种间的竞争情况,本研究中从蚊虫捕获数量上可以看出存在较明显的种间竞争。而新疆伊蚊与其他各蚊种间的生态位重叠指数均较低,表明新疆伊蚊对于栖息生境的要求与其他蚊种之间存在较大差异,成为新疆的特有蚊种。
系统聚类分析结果说明乌伦古河支流布尔根河湿地、额尔齐斯河下游湿地及伊犁河下游湿地的蚊类群落构成较为相似,艾比湖湿地和额敏河河口湿地蚊类群落构成较为相似。
本研究因只考虑了物种一个因素,未进行环境因素分析,因此采用了DCA排序分析。通常情况生态位较窄的物种多分布于排序图外围,生态位较宽的物种多位于排序图中心。本研究中生态位宽度较窄的新疆伊蚊、阿拉斯加脉毛蚊、哈萨克斯坦伊蚊等蚊种均位于排序图外围,而生态位宽度较宽的刺扰伊蚊、里海伊蚊、尖音库蚊、背点伊蚊、米赛按蚊等均位于排序图中心,白黑伊蚊、黄色伊蚊、环跗脉毛蚊以及哈萨克斯坦伊蚊与阿拉斯加脉毛蚊生态位重叠指数均为1.0000,在排序图中共享同一个位点。刺扰伊蚊、赫坎按蚊、凶小库蚊、里海伊蚊、尖音库蚊、米赛按蚊、背点伊蚊在排序图中相互间的距离相对较近,说明这类蚊种栖息生境很相似,从而构成了新疆北部干旱区湿地蚊类的常见种。
| [1] | 赵彤言,董言德,陆宝麟. 我国尖音库蚊复合组的聚类分析[J]. 寄生虫与医学昆虫学报,1999,6(1):39-45. |
| [2] | 张桂林,张建江,马德新. 新疆伊犁地区蚊虫调查及防治概况[J].中华卫生杀虫药械,2004,10(2):130-132. |
| [3] | 张桂林,刘斌,韩增宪. 蚊蚋对新疆某边防部队官兵健康危害的调查[J]. 中华卫生杀虫药械,2006,12(3):185-186. |
| [4] | 张桂林,郑重,张建江. 新疆北疆边境地区重要蚊虫及其防治[J].中华卫生杀虫药械,2008,14(5):417-418. |
| [5] | 张映梅,郭晓霞,赵彤言,等. 新疆哈巴河地区蚊种组成及昼夜消长规律调查研究[C]//中华预防医学会,中国疾病预防控制中心.第二届媒介生物可持续控制国际论坛论文集. 北京:中华预防医学会,中国疾病预防控制中心,2008:2. |
| [6] | 韩招久,王忠灿,王长军,等. 在某部营区利用CO2诱蚊器采样的物种优势度与多样性分析[J]. 中国媒介生物学及控制杂志, 2010,21(5):419-421. |
| [7] | 吕志,付士红,李铭华,等. 中国新疆部分地区蚊传黄病毒感染调查[J]. 疾病监测,2011,26(6):431-434. |
| [8] | 颜忠诚,安继尧,虞以新. 蚊虫地理生态位及其重叠群划分的初步研究[J]. 动物学研究,1997,18(3):293-297. |
| [9] | Hillebrand H. On the generality of the latitudinal diversity gradient[J]. Am Nat,2004,163(2):192-211. |
| [10] | Schäfer ML, Lundkvist E, Landin J, et al. Influence of landscape structure on mosquitoes (Diptera:Culicidae) and dytiscids (Coleoptera:Dytiscidae) at five spatial scales in Swedish wetlands[J]. Wetlands,2006,26(1):57-68. |
| [11] | Li L,Bian L, Yakob L, et al. Temporal and spatial staBility of Anopheles gamBiae larval haBitat distribution in Western Kenya highlands[J/OL]. Int J Health Geogr,2009,8:70-80[2014-02-26]. http://www.ij-healthgeographics.com/content/8/1/70. |
| [12] | Rueda1 ML, Brown TL, Kim HC,et al. Species composition, larval haBitats, seasonal occurrence and distribution of potential malaria vectors and associated species of Anopheles(Diptera: Culicidae) from the Republic of Korea[J/OL]. Malaria J,2010,9:55-65.[2014-02-06]. http://www.malariajournal.com/content/9/1/55. |
| [13] | Overgaard HJ, Ekbom B, Suwonkerd W, et al. Effect of landscape structure on anopheline mosquito density and diversity in northern Thailand: implications for malaria transmission and control[J]. Landscape Ecol,2003,18(6):605-619. |
| [14] | 葛军旗,孙肖红,龚正达,等. “三江并流”自然遗产地澜沧江流域居民区蚊类多样性的空间分布格局[J]. 生物多样性,2008,16(1):24-33. |
| [15] | 张菊仙,龚正达,李四全,等. “三江并流”自然遗产地怒江流域居民区蚊类多样性的空间分布格局[J]. 寄生虫与医学昆虫学报, 2008,15(4):213-222. |
| [16] | Ayala D, Costantini C, Ose K, et al. HaBitat suitaBility and ecological niche profile of major malaria vectors in Cameroon[J/OL]. Malaria J,2009,8:307-321.[2014-02-26]. http://www.malariajournal.com/content/8/1/307. |
| [17] | Cardo MV, Vezzani D, Carbajo AE. The role of the landscape in structuring immature mosquito assemblages in wetlands[J]. Wetlands Ecol Manage,2013,21:55-70. |
| [18] | 李洯,朱金兆,朱清科. 生态位理论及其测度研究进展[J]. 北京林业大学学报,2003,25(1):100-107. |
| [19] | 李斌,李素清,张金屯. 云顶山亚高山草甸优势种群生态位研究[J]. 草业学报,2010,19(1):6-13. |