 2014, Vol. 25
2014, Vol. 25服务
文章信息
- 彭真, 王勇, 范尊龙, 李亚衡, 徐云虎, 贺兵, 王玉梅, 赵景瑞
- PENG Zhen, WANG Yong, FAN Zun-long, LI Ya-heng, XU Yun-hu, HE Bing, WANG Yu-mei, ZHAO Jing-rui
- 内蒙古浑善达克地区黑线仓鼠的食性及其动态
- Food consumption of striped hamster (Cricetulus barabensis) and its dynamic changes in Hunshandake sandy land of Inner Mongolia, China
- 中国媒介生物学及控制杂志, 2014, 25(5): 408-412
- Chines Journal of Vector Biology and Control, 2014, 25(5): 408-412
- 10.11853/j.issn.1003.4692.2014.05.006
-
文章历史
- 收稿日期:2014-03-26




2 中国科学院大学,北京 100049;
3 内蒙古锡林郭勒盟镶黄旗草原工作站;
4 内蒙古锡林郭勒盟太仆寺旗气象局;
5 内蒙古锡林郭勒盟太仆寺旗土壤肥料工作站
2 University of Chinese Academy of Sciences, Beijing 100049,China;
3 Xianghuangqi Research Station of Grassland;
4 Taipusi Banner Meteorological Bureau, Inner MongoliaTaipusi Banner;
5 Taipusi Banner Agency for Soil and Fertilizer, Inner Mongolia Taipusi Banner
食物是动物生存和繁殖所需要的能量来源,一直 以来,食性研究都是生态学最为活跃的研究领域之 一[1]。对植食性动物的食性及其季节性研究,能够了 解动物的生活史、觅食对策等问题,对动植物系统协同 进化,植被演替等理论的发展有重要作用。前人对植 食性动物的食性进行了大量研究,广泛涉及到灵长 类[2]、鱼 类 [3,4]、鸟 类 [5]。 提出营养假说(nutritive hypothesis)[6],植物次生化合物假说(plant secondary compounds hypothesis)[7],营 养 平 衡 假 说(nutrient balance hypothesis)[8],最优觅食理论(optimal forging theory)[9],条件性气味回避性假说(conditional flavor aversion hypothesis)[10]。也有许多学者开展了啮齿动 物食性和食量的研究[11,12,13,14,15],20世纪90年代以来,频繁 的鼠害暴发啃食牧草和草场过度利用加剧了草地沙化,导致严重的草地资源损失[16]。因此,开展草原啮齿 动物的基础性食性研究,对于防控鼠害和保护草地资 源有重要意义。
黑线仓鼠(Cricetulus barabensis)为我国北方农田 和草地的主要害鼠之一,是内蒙古自治区锡林郭勒浑 善达克沙地的优势鼠种之一[17]。作为沙地生态系统中 的一部分,在能量流动、物质循环中发挥了重要作用。 黑线仓鼠主要取食植物种子,影响着沙丘植被的分布 与演替。此外,黑线仓鼠还是多种媒介蚤的宿主,可传 染多种人兽共患疾病[18,19,20]。因此,对黑线仓鼠的综合 治理研究,对抑制该地区沙化和防控鼠疫传播有重要 意义。对黑线仓鼠的研究大多集中在分子遗传以及种 群数量等方面[21,22,23,24,25,26],在食性动态方面报道较少,王淑卿 等[27]报道了河北省境内黑线仓鼠主要取食农作物种 子,同时还取食草籽和根、茎、叶、花、果实及动物性食 物。邢林和卢浩泉[28]报道了山东省境内笼养黑线仓鼠 喜食花生、大豆、玉米。然而对浑善达克沙地的黑线仓 鼠的食性还没有系统研究。为此,笔者将2003-2004 年在内蒙古浑善达克沙地捕获的黑线仓鼠进行分析, 为相关研究提供参考依据。 1 材料与方法 1.1 样地概况
研究地点为内蒙古锡林郭勒盟正蓝 旗乌日图苏木和阿巴嘎旗洪格尔苏木境内浑善达克沙 丘腹地(43°07′N,116°03′E)。年平均温度1.7℃,7 月平均气温22℃,1月平均气温-21℃,无霜期104 d, 多年平均年降雨量366.8 mm,多年平均蒸发量 1936.2 mm [29]。植被覆盖度多在12%以下,平均3%。
样区主要以固定沙丘为主,半流动性沙丘罕见,草场退 化 、沙化程度高 。主要植被为沙米(Agriophyllum squarrosum)、兴安虫实(Corispermum chinganicum)、沙 竹(Phyllostachys propinqua)、小叶锦鸡儿(Caragana microphylia)、狗 尾 草(Setaria viridis)、紫 花 苜 蓿 (Medicago satival)、杂花苜蓿(Melissitus ruthenica)、黄 柳 (Salix gordejevii) 、狭叶锦鸡儿 (Caragana stenophylla)、羊 柴(Hedysarum laeve)、山 天 冬 (Asparagus daurius)、叉分蓼(Polygonum divaricatum) 和猪毛菜(Salsola collina)等。主要鼠类为小毛足鼠 (Phodopus roborovskii)、三趾跳鼠(Dipus sagitta)、黑线 仓 鼠、五 趾 跳 鼠(Allactaga sibirica)、达 乌 尔 黄 鼠 (Spermophilus dauricus)、黑 线 毛 足 鼠(Phodopus sungorus)、棕 背鼠平(Myodes rufocanus)、莫 氏 田 鼠 (Microtus maximowizcii)、花鼠(Tamias sibiricus)、子午 沙 鼠(Meriones meridianus)、草 原 鼢 鼠(Myospalax aspalax)和鼹形田鼠(Ellobiustancreis)等 [30]。 1.2 数据采集
2003年开始对浑善达克沙地的乌日 图和洪格尔样地逐月夹捕取样,每月采样点6个,夹日 数为2400个,每日检查2次,每月连续检查3 d。为避 免连续夹捕对沙地鼠类种群数量的影响,每个采样点 只夹捕1次,然后进行食性数据的采集。2003年3-5 月受SARS的影响未能如期取样,因而本研究选择 2003年6-10月和2004年3-5月,该时间段内共捕获 黑线仓鼠169只。2003年11月至2004年2月,由于天 气严寒,未进行专门的夹线,数据来源于定位站北麓的 沙地观察结果[13]。 1.3 食性判定方法
对小型仓鼠的食性判定,参照张 新阶等[30]方法,以颊囊内储存的食物来判定其主要食 物名称,而食物主要类别(植物种子、植物茎叶、昆虫) 的比例构成则依赖胃内容物的检视结果。为定量表述 黑线仓鼠的食性变化,将捕获的黑线仓鼠颊囊内种子 组分的相对多度、出现频次以及种子的重要性进行 分级:
在种子的相对多度分级过程中先将种子体积相仿 的小种子作为基本单位,如杂花苜蓿、沙米、狗尾草、兴 安虫实等种子,小型种子1个为1单位;中型种子如沙 竹种子折算为5个单位,每个沙竹种子按照5个种子单 位计数;大型种子如小叶锦鸡儿种子、山天冬浆果折算 15个种子单位,按照这种折算方法,以颊囊内出现食 物的黑线仓鼠作为总样本,则可计算颊囊内各种植物 种子的平均数量,以此确定颊囊内各种种子的相对多 度:①1~10单位记为“1”;②11~50单位记为“2”;③ 50单位以上记为“3”。
种子的出现频率:在种子的出现频率分级过程中 将频次区分“有或无”,而不区分数目的多寡,这里的统 计样本是所有的个体,颊囊内未出现食物的黑线仓鼠 也计入统计样本。以此确定种子的出现频率:①<5% 记为“1”;②5%~15%记为“2”;③16%~30%记为“3”; ④30%以上记为“4”。 种子重要性:如果某种植物种子占颊囊中所有种 子的比例>20%,则定为主要食物,记为“3”;如果所占 比例在5%~20%则定为次要食物,记为“2”;<5%则 为一般食物,记为“1”。 2 结 果 2.1 食物组成
胃内容物的检视结果表明,浑善达克 沙地黑线仓鼠取食植物种子、植物茎叶和昆虫。其中 植物种子是黑线仓鼠主要食物类型 ,占取食的 55.0%~100%;昆虫的取食高峰在7月,约占取食的 8.9%;而植物茎叶的取食最高峰出现在5月,占食物的 40.0%。颊囊内植物种子鉴别结果表明,浑善达克沙地黑线仓鼠取食兴安虫实种子、沙米种子、狗尾草种 子、杂花苜蓿种子、小叶锦鸡儿种子、榆树(Ulmus pumila)种子。其中兴安虫实、狗尾草、小叶锦鸡儿在 黑线仓鼠取食的种子中相对多度、出现频率、重要度都 很高,其他几种种子相对较低;8月在部分个体中还发 现少量麦瓶草(Silene cnoidea)种子,9月发现部分个体 内出现叉分蓼种子和山天冬浆果,10月偶尔也发现沙 竹种子。 2.2 食物组成的动态变化
浑善达克沙地黑线仓鼠 每月的取食变化明显(图 1),3、4月取食基本是植物种 子,全年取食植物种子呈“V”形。4月以后天气变暖, 黑线仓鼠取食植物茎叶和昆虫的比率增加,尤其在5 月植物开始萌发,黑线仓鼠取食植物茎叶的比率为 40.0%。此时植物种子所占比率最低平均为58.8%。5 月之后,黑线仓鼠取食植物种子开始显著下降,仅占取 食的少部分。5月以后昆虫的活动开始频繁,黑线仓 鼠对昆虫的取食比率增加,直到7月平均占取食的 8.9%。之后比率开始下降。

|
| 图 1 黑线仓鼠胃内容物动态 Figure 1 Dynamics of foods in the stomach of C. barabensis |
通过Friedman秩和检验表明,黑线仓鼠取食种子 的相对多度在各月分布不同(P≤0.01)。兴安虫实种 子相对多度等级值只有在6、7月相对较低,而在6月后 狗尾草种子的值一直保持较高水平。小叶锦鸡儿种子 的最高值分布在7月,杂花苜蓿种子的最高值分布在7、 8月,其他植物种子的值在各月分布比较均匀(表 1)。
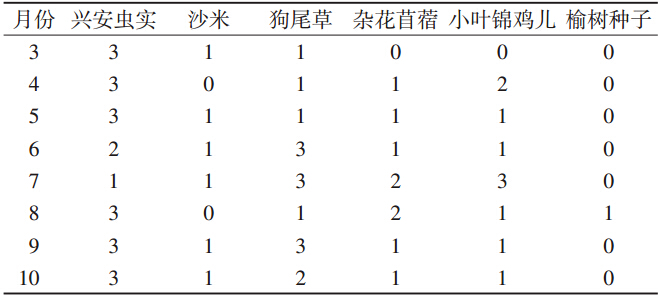 |
黑线仓鼠颊囊植物种子出现频次指数可以看出取 食某种子的样本在总体样本中所占比率的高低。黑线 仓鼠取食种子的样本数与总体样本比率值等级在各月 分布不同(Friedman秩和检验,P≤0.01)。兴安虫实出 现频率等级除6、7月外,一直处在最高位置。狗尾草 种子出现频率在6月后处在高水平值,小叶锦鸡儿种 子所占频率最高值分布在7月,杂花苜蓿种子最高值 分布在8月,其他种子的频率较低(表 2)。
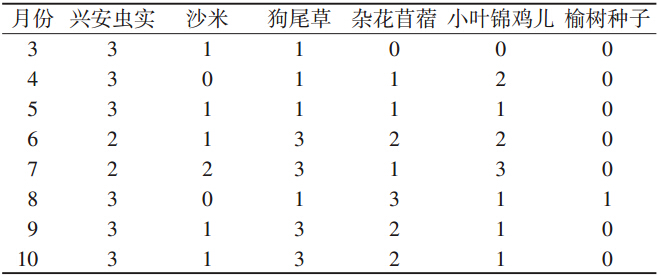 |
通过Friedman秩和检验表明,黑线仓鼠颊囊内种 子重要程度指数等级在各月分布不同(P≤0.01)。兴 安虫实种子重要程度指数值除6、7月外一直最高。6 月值最大的是狗尾草种子,7月是小叶锦鸡儿种子 (表 3)。
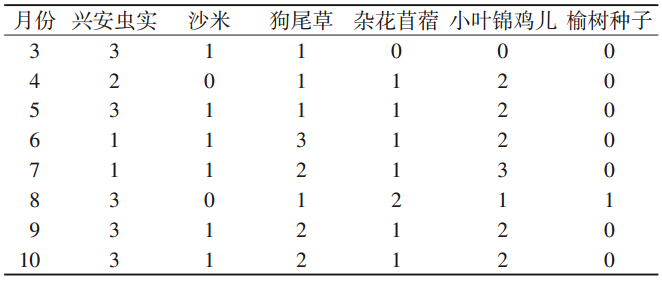 |
浑善达克沙地黑线仓鼠取食该地区的植物种子、 植物茎叶和昆虫,其中植物种子来自兴安虫实、沙米、 狗尾草、杂花苜蓿、小叶锦鸡儿和榆树。浑善达克沙地 沙化严重,兴安虫实、沙米和冷蒿为主要植被,盖度为 10.0%~30.0% [31]。以狗尾草为主的群落,狗尾草盖度 平均为10.5% [32]。而以小叶锦鸡儿为主的灌丛,小叶锦 鸡儿盖度为1.9%左右[33]。因而黑线仓鼠的食谱中,兴 安虫实种子、狗尾草种子、小叶锦鸡儿种子占主导地 位,是因为这几种植物种子在浑善达克沙地的库存量 比较丰富,并且其可觅食性和可搬运性程度较高。此与沙地中另外一种优势鼠种——小毛足鼠的食性[13]有 一定的差异,小毛足鼠很少取食兴安虫实种子和狗尾 草种子,尤其在食物充沛的7-9月,但黑线仓鼠很少 取食沙竹种子,可能与其攀援能力不如小毛足鼠有 关。这也反映出同域分布的2种仓鼠在食性上具有 明显的生态位分离,因而这2种鼠可以在沙地演替的 各个阶段,都能在沙地生境中稳定地分布,也可能表 明了黑线仓鼠对种子的特异选择性较弱,一般情况 下优先选择蕴藏量巨大的种子资源。另外浑善达克 沙地3-6月种子还没有成熟,可能是兴安虫实等种子 的库存量大,黑线仓鼠在几个月都是取食的前一年的 种子。除植物种子外,黑线仓鼠还取食昆虫食物和部 分植物幼苗及茎叶,可能是取食昆虫能够补充蛋白营 养,促进了黑线仓鼠的种群繁殖。取食部分植物幼苗 及茎叶是为了弥补食物的不足。此与小毛足鼠的生活 习性相近,这种食性的变化可视为对环境的长期适应 结果。
浑善达克沙地位于内蒙古高原,地处中纬度,属中 温带半干旱、大陆性季风气候,气候变化明显,降雨很 不稳定,蒸发量大[34]。植物多样性和茂盛度季节变化 明显,因而黑线仓鼠的食性也具有明显的季节性。3- 6月黑线仓鼠主要取食植物种子,但是取食的植物茎 叶和昆虫会增加。可能是该时间段植物种子都还未成 熟,是食物匮乏期,黑线仓鼠只能取食遗落的陈年种 子,为了生存和繁殖的需要,选择植物茎叶和蛋白质多 的昆虫。6月以后,黑线仓鼠取食种子的比率上升,可 能是各种植物种子相继成熟,黑线仓鼠倾向于选择可 搬运、易储存的植物种子,为度过气候条件恶劣的冬季 做准备。7月和8月黑线仓鼠取食昆虫的比率较多,这 是由于在内蒙古地区7、8月是昆虫活动和繁殖的高峰 期;黑线仓鼠每月取食植物种子的种类及多少也有不 同。在沙地植物中,兴安虫实覆盖度很高,种子库相对 其他植物丰富,因此成为黑线仓鼠首选的食物,6月以 后,黑线仓鼠也会大量取食狗尾草、小叶锦鸡儿种子, 是由于这些种子相继成熟,黑线仓鼠会在可取食性和 可利用性上选择最优的觅食策略。2003年11月至 2004年2月,沙地夜间已非常寒冷,并有积雪覆盖,未 进行夹线调查,因此冬季的食性未知,但从沙地黑线仓 鼠的足迹痕迹来分析,黑线仓鼠冬天在兴安虫实、沙米 等植株下停留,采集植株上残余的种子。地面上的取 食也可能是作为越冬食物的补充来源之一。
鼠类作为生态系统中的一个重要功能群,对生境 植被的更新起重要作用。一方面,鼠类暴发对植物种 子有很大的取食压力,不利于植被的更新[35];另一方 面,许多植物的更新又必须依赖鼠类对种子的散布和 埋藏,对植物更新是有利的[36,37]。通过对黑线仓鼠食 性的季节变化分析可以看出,黑线仓鼠主要取食沙地 植物种子,还取食少量的植物茎叶,这对沙地植物种子 库和植物营养生长的破坏作用极大。黑线仓鼠这种取 食作用对于植被稀疏的沙地生境来说,严重影响了其 植被的恢复与自我更新进程。同时,黑线仓鼠还具有 搬运和存储食物的习性。它将小叶锦鸡儿种子、狗尾 草种子和兴安虫实种子等食物充塞颊囊转运的过程, 其实也是沙地植物种子传播的过程。并且有研究表明 植物种子储存在土壤中能提高萌发率[38]。尤其对于沙 丘的演替后期,沙丘植被茂盛之后,黑线仓鼠取食的那 些种子只占沙地植被种子产量很小的一部分。从这个 角度来说,黑线仓鼠对沙地植被演替具有重要作用。
志谢 中国科学院动物研究所宛新荣副研究员对实验和论文 提出宝贵意见,内蒙古草原生态系统定位研究站和内蒙古草原 动物生态研究站对野外实验的开展提供了很大的平台,一并志谢。
| [1] | Wang SW,Macdonald DW. Feeding habits andniche partitioning in a predator guild composed of tigers,leopards and dholes in a temperate ecosystem in central Bhutan[J]. J Zool,2009,277(4): 275-283. |
| [2] | 黄中豪,黄乘明,周岐海,等.黑叶猴食物组成及其季节性变化[J]. 生态学报,2010,30(20):5501-5508. |
| [3] | 郭爱,周永东,金海卫,等. 东海黄鲫的食物组成和食性的季节变 化[J].现代渔业信息,2010,25(8):10-13,27. |
| [4] | 杨学峰,谢从新,马宝珊,等. 拉萨裸裂尻鱼的食性[J]. 淡水渔业,2011,41(4):40-44,49. |
| [5] | 张树舜,张淑兰. 长耳鸮食性的初步分析[J]. 动物学杂志,1990, 25(5):23. [6]Nolte DL,Provenza FD. Food preferences in lambs after exposure to flavors in solid foods[J]. Appl Anim Behav Sci,1992,32(4): 337-347. |
| [7] | Torregrossa AM,Dearing MD. Nutritional toxicology of mammals: regulated intake of plant secondary compounds[J]. Funct Ecol, 2009,23(1):48-56. |
| [8] | Yasmin JC.Arabidopsis thalianaresistance to insects,mediated by an earthworm?produced organic soil amendment[J]. Pest Manag Sci,2011,67(2):233-238. |
| [9] | Skorka P,Lenda M,Martyka R,et al. The use of metapopulation and optimal foraging theories to predict movement and foraging decisions of mobile animals in heterogeneous landscapes[J]. Landscape Ecol,2009,24(5):599-609. |
| [10] | Roman C,Lin JY,Reilly S. Conditioned taste aversion and latent inhibition following extensive taste preexposure in rats with insular cortex lesions[J]. Brain Res,2009,1259:68-73. |
| [11] | 冯志勇,邱俊荣,姚丹丹,等. 自然条件下板齿鼠的食物结构研究[J].中国媒介生物学及控制杂志,2010,21(1):23-25. |
| [12] | 何晓瑞. 我国小竹鼠洞系结构和食性的研究[J]. 动物学杂志, 1994,29(1):35-37. |
| [13] | 宛新荣,刘伟,王广和,等. 浑善达克沙地小毛足鼠的食量与食性动态[J].生态学杂志,2007,26(2):223-227. |
| [14] | 吴林,张美文,李波. 洞庭湖区东方田鼠的食物组成调查[J]. 兽类学报,1998,18(4):43-52. |
| [15] | 雍仲禹,张美文,郭聪,等. 洞庭湖区黑线姬鼠食性调查[J]. 动物学杂志,2012,47(3):115-121. |
| [16] | 钟文勤,樊乃昌.我国草地鼠害的发生原因及其生态治理对策[J]. 生物学通报,2002,37(7):1-4. |
| [17] | 武晓东,付和平. 内蒙古半荒漠与荒漠区的啮齿动物群落[J]. 动物学报,2005,51(6):961-972. |
| [18] | 杜国义,史献明,王海峰,等. 河北省鼠疫自然疫源地黑线仓鼠寄生蚤的调查[J].中国媒介生物学及控制杂志,2008,19(2):157-158. |
| [19] | 孙养信,阮春来,白江春,等. 陕西省鼠疫疫区夜行鼠寄生蚤调 查[J]. 中国病原生物学杂志,2008,3(2):133-135. |
| [20] | 王峰,任清明,刘国平,等. 吉林省洮南地区鼠类及鼠疫血清学调查[J].中国媒介生物学及控制杂志,2007,18(6):501-502. |
| [21] | Song ZG,Wang DH. The maximum metabolizable energy intake and the relationship with basal metabolic rate in the striped hamster Cricetulus barabensis[J]. Acta Theriol,2002,47(4):417-423. |
| [22] | Yan CL,Xu T,Qu X,et al. Agricultural irrigation mediates climatic effects and density dependence in population dynamics of Chinese striped hamster in North China Plain[J]. J Anim Ecol, 2013,82(2):334-344. |
| [23] | 董维惠,侯希贤,林小泉,等.黑线仓鼠种群数量动态预测研究[J]. 生态学报,1993,13(4):300-305. |
| [24] | 董维惠,侯希贤,杨玉平,等. 草原和农田几种主要鼠种数量动态 研究及预测[J].中国草地学报,2008,30(5):90-95. |
| [25] | 武文华,付和平,武晓东,等. 应用马尔可夫链模型预测长爪沙鼠 和黑线仓鼠种群数量[J].动物学杂志,2007,42(6):69-78. |
| [26] | 张琪,薛慧良,徐金会,等. 黑线仓鼠CRH基因的编码序列及系 统进化分析[J].生物技术通报,2012(10):88-94. |
| [27] | 王淑卿,杨荷芳,郝守身,等. 黑线仓鼠的食性与食量[J]. 动物学报,1992,38(2):156-164. |
| [28] | 邢林,卢浩泉. 黑线仓鼠的食性及防治阈值的探讨[J]. 动物学杂志,1990,25(4):29-33. |
| [29] | 丁国栋,李素艳,蔡京艳,等. 浑善达克沙地草场资源评价与载畜 量研究:以内蒙古正蓝旗沙地为例[J]. 生态学杂志,2005,24 (9):1038-1042. |
| [30] | 张新阶,王广和,刘伟,等. 浑善达克沙地三趾跳鼠的食性与繁殖 特征的初步分析[J].动物学杂志,2007,42(3):9-13. |
| [31] | 刘树林,王涛. 浑善达克沙地的土地沙漠化过程研究[J]. 中国沙漠,2007,27(5):719-724. |
| [32] | 宋创业,郭柯. 浑善达克沙地中部丘间低地植物群落分布与土壤 环境关系[J].植物生态学报,2007,31(1):40-49. |
| [33] | 初玉,杨慧玲,朱选伟,等. 浑善达克沙地小叶锦鸡儿灌丛的空间 异质性[J].生态学报,2005,25(12):3294-3300. |
| [34] | 宝玉. 对浑善达克沙地荒漠化现状及其气候的影响简析[J]. 北 方经济,2012(19):64-66. |
| [35] | 秦姣,施大钊,贠旭疆,等. 模拟条件下布氏田鼠对草原植被的作用 2.鼠密度对植物群落结构及根系的影响[J]. 草业科学, 2009,26(7):127-132. |
| [36] | Sanguinetti J,Kitzberger T. Factors controlling seed predation by rodents and non?nativeSus scrofain Araucaria araucanaforests: potential effects on seedling establishment[J]. Biol Invasions, 2010,12(3):689-706. |
| [37] | Birkedal M,Fischer A,Karlsson M,et al. Rodent impact on establishment of direct?seeded Fagus sylvatica,Quercus robur and Quercus petraeaon forest land[J]. Scand J Forest Res,2009,24 (4):298-307. |
| [38] | 张知彬,王福生. 鼠类对山杏种子存活和萌发的影响[J]. 生态学报,2001,21(11):1761-1768. |