 2024, Vol. 35
2024, Vol. 35扩展功能
文章信息
- 刘全超, 邹亚明, 兰策介
- LIU Quan-chao, ZOU Ya-ming, LAN Ce-jie
- 3种蚊虫优势度计算方法比较研究
- Comparative study on three calculation methods for mosquito dominance
- 中国媒介生物学及控制杂志, 2024, 35(3): 329-333
- Chin J Vector Biol & Control, 2024, 35(3): 329-333
- 10.11853/j.issn.1003.8280.2024.03.013
-
文章历史
- 收稿日期: 2023-12-25



物种优势度是指在一个生物群落中某种物种的地位与作用,它被用来量化一个物种子集对一个群落总丰度的支配程度[1]。研究群落的物种优势度有助于确定群落中物种分布的均匀程度、确定优势物种,在群落生态学研究和物种多样性保护方面应用广泛。物种优势度主要体现在物种的个体数量、生物量、体积、出现频率等方面,由于生存状态、活动类型等特征的差异,不同物种计算优势度的方法不同。传统的计算方法主要依赖于个体数以及出现频率等变量,如Simpson、Lloyd和Austin指数等是基于群落中物种的个体数量和物种数量来计算物种优势度[2-4];McNaughton指数则是根据物种的总体出现频率及个体数量来确定浮游生物的优势种,目前也有较多应用[5]。从生态系统物质循环和能量流动的角度分析,物种优势度是指某一物种在群落中相对于其他物种对所处生态位的物质和能量流动影响的强弱程度。这包含3个维度的影响,一是目前该物种所持有的物质和能量总和在群落中的占比,二是在时间尺度上该物种对生态位的影响尺度,三是在空间尺度上该物种对生态位的影响尺度。由于物种分布的时空异质性,在全面考虑这3个影响维度的情况下,才能更完整地反映物种优势度。
蚊虫是重要的医学昆虫,不同的蚊虫传播不同的疾病,如按蚊(Anopheles)传播疟疾、伊蚊(Aedes)传播寨卡病毒病和登革热、库蚊(Culex)传播流行性乙型脑炎等[6-9]。通常情况下媒介蚊虫处于优势种时才会造成传染病的广泛流行[10-11]。因此,评估蚊虫优势度对评估一定生态位内潜在蚊媒传染病流行风险的时空分布具有很强的预警作用。由于不同蚊种的时间、空间分布的差异性,目前在计算蚊种优势度时,会出现个别点位的单个蚊种密度远远高于其他所有点位的蚊种密度之和,虽然个别点位的影响范围非常有限,但数量又处于绝对优势,最后导致调查结果和区域整体实际情况不一致[12]。因此本文从生物量、时间尺度和空间尺度3个维度探讨优势度的计算方法,提出了Time-Space指数,并与上述Berger-Parker指数和McNaughton指数的统计结果进行比较分析,为进一步了解和优化物种优势度的测度提供参考。
1 材料与方法 1.1 数据来源 1.1.1 蚊虫调查本研究资料来源于江苏省无锡市2012-2021年蚊虫监测数据。其中2012-2015年设置居民区、公园、医院各5个点位,农户、牲畜棚各4个点位,2016年各生境分别设置5个点位,2017-2021各生境分别设置4个点位。在每年3-11月的各月使用诱蚊灯(功夫小帅牌)法开展2次蚊虫调查,调查时间为日落前1 h至次日日出后1 h,每2次监测间隔时间 > 10 d。蚊密度计算公式为:D=X/(L×h),其中,D为蚊密度、X为捕获雌蚊数、L为诱蚊灯数、h为诱蚊时间。
1.1.2 蚊虫标本处理将捕获的蚊虫进行分类、计数,将蚊虫按种类分组,每组100只,烘干至充分脱水后称重并记录(连续3次过夜烘干后称量,质量无变化,视为充分脱水),计算不同蚊种充分脱水后单只蚊虫的平均干重,记作生物量。
1.2 优势度的测度方法 1.2.1 Berger-Parker指数Berger-Parker指数由Berger和Parker于1970年提出,是在物种个体数量上,描述一个物种在群落中分布的优势程度,是目前最为常用的优势度计算方法。其计算公式为:
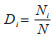
|
式中,Di为物种i的相对优势度,Ni为物种i的个体数量,N为全部物种的个体总数。
1.2.2 McNaughton指数Berger-Parker指数的计算主要依托于生物的个体数量,McNaughton指数则是在传统的基础上新引入了频率变量,主要根据物种的出现频率和个体数量来确定优势种,其计算公式为:
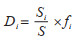
|
式中,Di为物种i的相对优势度,Si为第i个物种的个体数量,S为所有物种的总个体数,fi为第i个物种在各点位出现的频率。
1.2.3 Time-Space指数Time-Space为本文提出物种优势度的测度方法,是在上述2种以个体数量为依托的传统计算方法的基础上,引入时间和空间变量,提出Time-Space指数,计算公式为:
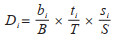
|
式中,Di为物种i的相对优势度,取值范围为(0,1],当为1时,表明目标生态位中某一物种对该生态位的物质与能量流动在生物方面的影响为100%。B是群落中的所有物种的总生物量,根据蚊虫的总数及质量确定,bi表示i蚊种的生物量。T是时间总周期,根据研究的时间尺度确定具体周期的长短,ti表示物种在总周期内出现的时间频率。S为空间总盖度,根据生态位的总面积为目标区域的空间总盖度,si表示物种在该区域出现的盖度总和。以年为周期统计每年各蚊种的个体数占比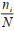
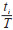
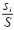
采用Excel 2019软件对数据进行统计分析。首先应用上述3种物种优势度的计算方法分别计算不同年份常见蚊种的优势度指数,再利用Kendall’s W系数评价3种指数的一致性,最后以不同年份的数据为重复,采用非参数Kruskal-Wallis H检验对不同指数进行差异性分析,P < 0.05为差异有统计学意义。
2 结果 2.1 蚊类的组成及其时空分布2012-2021年各年设置监测点位数量20~25个不等,时间断面数(即总采集次数)为每年18次,共捕获蚊虫27 281只,平均蚊密度为0.58只/(灯·h)。蚊虫种类主要为淡色/致倦库蚊(Cx. pipiens pallens/quinquefasciatus)、三带喙库蚊(Cx. tritaeniorhynchus)、白纹伊蚊(Ae. albopictus)、中华按蚊(An. sinensis)以及骚扰阿蚊(Armigeres subalbatus),平均干重分别为0.67、0.60、0.42、0.84和1.57 mg/只,其他蚊种数量较少未统计。淡色/致倦库蚊空间平均出现频率比为97.70%,时间出现频率比为94.4%,历年蚊虫监测结果见表 1。

|
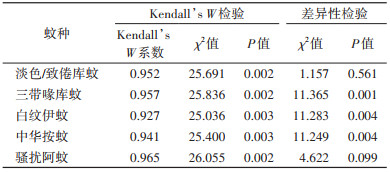
所有蚊种的Kendall’s W一致性系数均 > 0.900,三带喙库蚊、白纹伊蚊及中华按蚊的3种优势度指数差异均有统计学意义(χ2=11.365,P=0.001;χ2=11.283,P=0.004;χ2=11.249,P=0.004)。三带喙库蚊、白纹伊蚊和中华按蚊的Time-Space指数明显低于Berger-Parker指数(Z=-2.192,P=0.028;Z=-2.343,P=0.019;Z=-2.041,P=0.041),中华按蚊的McNaughton指数显著低于Berger-Parker指数(Z=-2.192,P=0.028);淡色/致倦库蚊与骚扰阿蚊的3种优势度指数差异无统计学意义(χ2=1.157,P=0.561;χ2=4.622,P=0.099)。各蚊种的3种优势度指数比较及检验见图 1、表 2。

|
| 图 1 江苏省无锡市2012-2021年5种常见蚊种的3种优势度指数比较 Figure 1 Comparison of three dominance indices of five common mosquito species in Wuxi, Jiangsu Province, 2012-2021 |
| |
|
不同蚊种的Kendall’s W一致性系数总体显著性均 < 0.05,说明Time-Space指数与McNaughton指数、Berger-Parker指数这2种目前主流的优势度计算方法整体具有一致性。淡色/致倦库蚊3种优势度的计算结果差异均无统计学意义,可能因为时空分布相对均匀。3种优势度指数的差异性结果显示,中华按蚊的McNaughton指数明显低于Berger-Parker指数,可能是因为将频率因素分开后,提高了分布异质性的分辨度;而三带喙库蚊、白纹伊蚊和中华按蚊的Time-Space指数明显低于McNaughton指数,说明在考虑空间分布及生物量因素后,分辨度进一步提高。
在现场群落生态学调查中往往通过设置采样点位,并以点位的调查信息来反映整体的概况。Berger-Parker指数和McNaughton指数作为传统的计算方法已有较多应用并得到广泛认同。例如黄海涛等[13]、杨立军和张丹丹[14]使用Berger-Parker指数研究了蝶类、蛾类等物种的多样性;刘潇等[5]在研究荣成近岸养殖海域浮游生物群落结构中使用了McNaughton指数计算物种优势度。本研究综合了物种个体、时间和空间3个维度提出了Time-Space指数,并在无锡市蚊虫群落结构研究中进行了验证,其对于三带喙库蚊、白纹伊蚊和中华按蚊的优势度结果明显低于Berger-Parker指数,提高了分辨优势物种的特异性。
蚊虫的分布受生态环境、温度、习性等多重因素的影响[15],个别蚊种例如三带喙库蚊、白纹伊蚊及中华按蚊等的分布具有聚集性,属偏态分布,白纹伊蚊成蚊的分布服从负二项分布[16],本次研究结果表明,Time-Space指数更加适用于此类时空异质性较高的物种分布特征研究,能更精细地反映其真实的优势度指数。Time-Space指数在应用的过程中要根据研究物种的习性对调查时间和调查点位进行设置,对于季节消长明显的物种要适当调整调查频率,调查点位间距设置也要考虑物种的活动范围等因素。Time-Space指数在应用过程中也存在一些问题,不同监测工具对不同蚊种的捕获能力有差异,本研究采用较为广谱的诱蚊灯法,其对于白纹伊蚊的诱捕能力较弱,因此结果可能会出现偏倚,另外调查点位设置的均衡性也会影响指数的结果。
Time-Space指数体现了物种时空分布异质性对物种优势度的影响,统计分析结果显示该指数提高了部分蚊种分布异质性的分辨度和反映了不同蚊种间相对优势度的具体状态。该结果也为其他物种优势度的评估提供了参考。
利益冲突 无
| [1] |
Fung T, Villain L, Chisholm RA. Analytical formulae for computing dominance from species-abundance distributions[J]. J Theor Biol, 2015, 386: 147-158. DOI:10.1016/j.jtbi.2015.09.011 |
| [2] |
Simpson EH. Measurement of diversity[J]. Nature, 1949, 163(4148): 688. DOI:10.1038/163688a0 |
| [3] |
Ma ZS, Ellison AM. A unified concept of dominance applicable at both community and species scales[J]. Ecosphere, 2018, 9(11): e02477. DOI:10.1002/ecs2.2477 |
| [4] |
Austin MP. Relationships among functional properties of Californian grassland[J]. Nature, 1968, 217(5134): 1163. DOI:10.1038/2171163a0 |
| [5] |
刘潇, 潘玉龙, 孙蓓蓓, 等. 荣成近岸养殖海域浮游动物群落结构及与环境因子的关系[J]. 现代农业科技, 2019(10): 177-179, 182. Liu X, Pan YL, Sun BB, et al. Structure of zooplankton community and its relation with environmental factors in Rongcheng adjacent waters[J]. Mod Agric Sci Tech, 2019(10): 177-179, 182. DOI:10.3969/j.issn.1007-5739.2019.10.113 |
| [6] |
汪圣强, 杨蒙蒙, 朱国鼎, 等. "一带一路"倡议下输入性蚊媒传染病的防控[J]. 中国血吸虫病防治杂志, 2018, 30(1): 9-13. Wang SQ, Yang MM, Zhu GD, et al. Control of imported mosquito-borne diseases under the Belt and Road Initiative[J]. Chin J Schisto Control, 2018, 30(1): 9-13. DOI:10.16250/j.32.1374.2017208 |
| [7] |
吴云姣, 范智玉, 刘桂芳, 等. 山东省2020-2021年蚊虫中乙型脑炎病毒感染率及系统进化分析[J]. 中国病毒病杂志, 2023, 13(1): 37-42. Wu YJ, Fan ZY, Liu GF, et al. Infection rate and phylogenetic analysis of Japanese encephalitis virus in mosquitoes in Shandong Province, 2020-2021[J]. Chin J Viral Dis, 2023, 13(1): 37-42. DOI:10.16505/j.2095-0136.2023.0008 |
| [8] |
Talapko J, Škrlec I, Alebić T, et al. Malaria: The past and the present[J]. Microorganisms, 2019, 7(6): 179. DOI:10.3390/microorganisms7060179 |
| [9] |
Monteiro FJC, Mourão FRP, Ribeiro ESD, et al. Prevalence of Dengue, Zika and Chikungunya viruses in Aedes (Stegomyia) aegypti (Diptera: Culicidae) in a medium-sized city, Amazon, Brazil[J]. Rev Inst Med Trop Sã o Paulo, 2020, 62: e10. DOI:10.1590/S1678-9946202062010 |
| [10] |
Odero JO, Nambunga IH, Wangrawa DW, et al. Advances in the genetic characterization of the malaria vector, Anopheles funestus, and implications for improved surveillance and control[J]. Malar J, 2023, 22(1): 230. DOI:10.1186/s12936-023-04662-8 |
| [11] |
刘起勇. 气候变化对中国媒介生物传染病的影响及应对: 重大研究发现及未来研究建议[J]. 中国媒介生物学及控制杂志, 2021, 32(1): 1-11. Liu QY. Impact of climate change on vector-borne diseases and related response strategies in China: Major research findings and recommendations for future research[J]. Chin J Vector Biol Control, 2021, 32(1): 1-11. DOI:10.11853/j.issn.1003.8280.2021.01.001 |
| [12] |
沈元, 孙俊, 兰策介, 等. 不同土地利用方式对蚊虫群落的影响[J]. 生态学杂志, 2012, 31(7): 1751-1755. Shen Y, Sun J, Lan CJ, et al. Effects of different land use types on mosquito community[J]. Chin J Ecol, 2012, 31(7): 1751-1755. DOI:10.13292/j.1000-4890.2012.0292 |
| [13] |
黄海涛, 李秋剑, 李志锐, 等. 澳门蝶类物种多样性及其区系研究[J]. 环境昆虫学报, 2016, 38(1): 87-94. Huang HT, Li QJ, Li ZR, et al. Study on species diversity and fauna of butterflies in Macao[J]. J Environ Entomol, 2016, 38(1): 87-94. DOI:10.3969/j.issn.1674-0858.2016.01.11 |
| [14] |
杨立军, 张丹丹. 井冈山自然保护区蛾类多样性及人为干扰的影响[J]. 环境昆虫学报, 2014, 36(5): 679-686. Yang LJ, Zhang DD. Diversity of moth communities and variation along artificial disturbance gradient in Mount Jinggangshan National Nature Reserve, China[J]. J Environ Entomol, 2014, 36(5): 679-686. DOI:10.3969/j.issn.1674-0858.2014.05.4 |
| [15] |
刘全超, 朱丁, 邹亚明, 等. 江苏省无锡市2012-2021年蚊虫生态学监测结果分析[J]. 中国媒介生物学及控制杂志, 2023, 34(4): 513-517. Liu QC, Zhu D, Zou YM, et al. An analysis of mosquito ecological surveillance results in Wuxi, Jiangsu Province, China, 2012-2021[J]. Chin J Vector Biol Control, 2023, 34(4): 513-517. DOI:10.11853/j.issn.1003.8280.2023.04.013 |
| [16] |
许峰, 王唐, 宋灿磊, 等. 诱蚊诱卵器捕获白纹伊蚊成蚊聚集趋势及分布型的研究[J]. 中国媒介生物学及控制杂志, 2021, 32(4): 481-486. Xu F, Wang T, Song CL, et al. Aggregation tendency and distribution pattern of adult Aedes albopictus mosquitoes captured by mosquito ovitraps[J]. Chin J Vector Biol Control, 2021, 32(4): 481-486. DOI:10.11853/j.issn.1003.8280.2021.04.020 |