 2024, Vol. 35
2024, Vol. 35扩展功能
文章信息
- 尚猛, 王璐, 姜宛均, 王晓旭, 冀好强, 梁莹, 刘起勇
- SHANG Meng, WANG Lu, JIANG Wan-jun, WANG Xiao-xu, JI Hao-qiang, LIANG Ying, LIU Qi-yong
- 2013-2021年气象因素及其交互作用对内蒙古自治区鄂尔多斯市鼠密度的影响
- Individual and interactive effects of meteorological factors on rodent density in Ordos, Inner Mongolia Autonomous Region, China, 2013-2021
- 中国媒介生物学及控制杂志, 2024, 35(2): 205-211
- Chin J Vector Biol & Control, 2024, 35(2): 205-211
- 10.11853/j.issn.1003.8280.2024.02.014
-
文章历史
- 收稿日期: 2023-08-31



2 传染病溯源预警与智能决策全国重点实验室, 中国疾病预防控制中心传染病预防控制所媒介生物控制室, 世界卫生组织媒介生物监测与管理合作中心, 北京 102206
2 National Key Laboratory of Intelligent Tracking and Forecasting for Infectious Diseases, Department of Vector Biology and Control, National Institute for Communicable Disease Control and Prevention, Chinese Center for Disease Control and Prevention, WHO Collaberating Centre for Vector Surveillance and Management, Beijing 102206, China
鼠类是哺乳类啮齿目动物的统称,作为多种病原体的贮存宿主和媒介,其种群密度的变化会对鼠疫、流行性出血热、钩端螺旋体病等鼠源疾病产生影响[1-3]。近年来,随着气候变化和极端天气事件频发,使得适宜鼠类生长繁殖的环境在时空范围上发生改变,导致鼠类的分布区域增加和年内危害期延长,进一步加剧了鼠源疾病的潜在风险。以黄胸鼠(Rattus tanezumi)为例,气候变暖使得其携带鼠疫耶尔森菌(Yersinia pestis,鼠疫菌)的活动范围扩展到北纬17°以南地区[4]。2010-2018年,人间鼠疫病例主要在中国西北部分省份零星报告,而近5年全国已报告鼠疫病例14例,其中鄂尔多斯市已连续2年报告人间鼠疫病例[5]。鄂尔多斯市处于蒙古高原荒漠草原长爪沙鼠(Meriones unguiculatus)自然疫源地[6],其四季分明的温带大陆性气候和连绵起伏的高平原为多种鼠类的生存提供了适宜的条件。因此,对当地鼠类进行系统、科学的生态学监测和密度数据分析,将有助于制定有针对性的鼠源疾病防控策略。
近年来,关于鼠密度的多项流行病学研究强调了气象因素在这一领域的重要性[7-8],但相关研究结果存在差异。关于温度的影响,多项研究发现在一定范围内,平均气温与鼠密度呈正相关[9-10];而在阿拉善荒漠地区的一项研究发现,温度升高抑制子午沙鼠(M. meridianus)种群数量增多[11]。鼠密度受气象因素的影响通常取决于各因素综合作用[12],然而,目前大多数研究仅限于独立考察各气象因素的效应[9, 13-16]。同时考虑到地区间自然生态、社会环境等存在较大差异,本研究利用鄂尔多斯市鼠密度的监测资料,采用广义可加模型(generalized additive model,GAM)分析气象因素对鼠密度的影响,并进一步研究气象因素间的交互作用对鼠密度的影响。
1 材料与方法 1.1 鼠密度资料监测数据来源于“中国疾病预防控制信息系统”的“鼠疫防治管理系统”模块,包含2013-2021年鄂尔多斯市鄂托克旗、鄂托克前旗、杭锦旗和乌审旗4个监测点的数据。监测数据包括监测月的布笼总数(个)、捕鼠总数(只)及各鼠种的捕获情况。在本研究中,鼠密度=捕鼠数/布笼总数×100%。
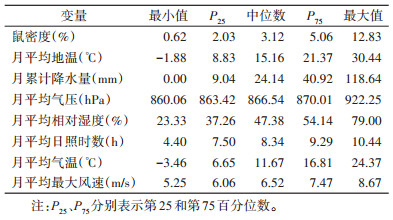
1.2 气象资料鼠类监测期间鄂尔多斯市同期气象资料来自中国科学院资源环境科学与数据中心(http://www.resdc.cn/Default.aspx),包括月平均地温(℃)、月累计降水量(mm)、月平均气压(hPa)、月平均相对湿度(%)、月平均日照时数(h)、月平均气温(℃)、月平均最大风速(m/s)7个气象因素。
1.3 统计学分析 1.3.1 描述性分析构建鄂尔多斯市鼠密度和监测期间气象数据的时间序列,采用四分位数描述其一般特征。
1.3.2 相关及滞后效应分析计算各气象因素之间及其与鼠密度的Spearman秩相关系数,以避免引入过多因素导致的共线性问题。结合Spearman秩相关系数,并考虑生物学合理性,仅选择保留主要的气象因素进行深入研究。由于气象因素的滞后效应,本研究选取最大滞后期为2个月,对主要气象因素进行滞后调整,包括滞后0月(lag0)、滞后1月(lag1)和滞后2月(lag2)。根据相关系数值的大小,确定各主要气象因素的最佳滞后期。
1.3.3 气象因素对鼠密度的影响鼠密度与气象因素的多因素分析采用类泊松回归的GAM[17],将各时点鼠密度变换后放入模型,鉴于气象因素对鼠密度的影响通常呈现非线性趋势[18],故在本研究中采用平滑样条的方式将各气象因素引入模型。综合模型的决定系数(R2)、偏差解释(deviance explained,单位为%)和广义交叉验证(generalized cross validation,GCV)选择最优模型。其最终公式如下:
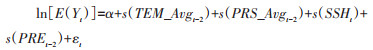
|
式中,E(Yt)代表第t监测月(鼠密度)的期望值,α为模型截距,TEM_Avg、PRS_Avg、SSH、PRE分别代表月平均气温、月平均气压、月平均日照时数、月累计降水量,s()代表平滑样条函数,εt是随机误差项。
1.3.4 交互作用分析依次运用GAMs进行分析,使用张量积平滑函数te()拟合主要气象因素的交互作用对鼠密度的影响,结果用平滑曲面展示。基本模型如下:
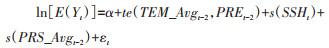
|
研究采用R 4.2.2软件中的“mgcv”程序包,取双侧检验,检验水准α=0.05。
2 结果 2.1 鄂尔多斯市鼠密度及月气象因素概况监测期间,在鄂尔多斯市总布笼83 192个,共捕鼠3 331只,总捕获率为4.00%。月平均气温中位数为11.67 ℃,月平均气压中位数为866.54 hPa,月平均日照时数中位数为8.34 h(表 1)。2013-2021年鄂尔多斯市总鼠密度及各鼠种密度时序图见图 1,尽管在2018年有个小高峰,但经Mann-Kendall趋势检验,总鼠密度呈现下降趋势[斜率(S)=-0.222,Z=-1.981,P=0.048),差异有统计学意义。其中子午沙鼠占比最高,为31.27%,近年来该鼠种密度逐渐下降(S=-0.086,Z=-1.147,P=0.252),而长爪沙鼠密度有所升高(S=0.041,Z=1.160,P=0.246),但二者趋势差异不明显。
|

|
| 图 1 内蒙古自治区鄂尔多斯市2013-2021年总鼠密度及各鼠种密度时序图 Figure 1 Time series diagram of total rodent density and rodent species density in Ordos, Inner Mongolia Autonomous Region, 2013 to 2021 |
| |
Spearman秩相关分析结果显示,鼠密度与月平均日照时数(rs=0.63)、月平均最大风速(rs=0.46)、月平均地温(rs=0.33)、月平均气温(rs=0.30)呈正相关(P < 0.05);而与月平均相对湿度(rs=-0.59,P < 0.05)、月平均气压(rs=-0.42,P < 0.05)、月累计降水量(rs < -0.01,P > 0.05)呈负相关(图 2)。考虑到既往研究均提示降水量对鼠密度存在密切关系[19],本研究结合生物学经验,最终将月平均日照时数、月平均气温、月平均气压和月累计降水量纳入模型,而与月平均日照时数密切相关的月平均最大风速、月平均相对湿度,与月平均气压相关的月平均地温不纳入模型,以避免共线性问题。

|
| 注:图中蓝点表示正相关,红点表示负相关,圆的大小和颜色深浅表示相关性的强弱。 图 2 内蒙古自治区鄂尔多斯市2013-2021年鼠密度和气象因素间的Spearman相关性分析 Figure 2 Spearman correlation analysis between rodent density and meteorological factors in Ordos, Inner Mongolia Autonomous Region, 2013-2021 |
| |
滞后分析显示,月平均气温、月累计降水量在滞后2月时与鼠密度相关性最高,月平均日照时数、月平均气压在滞后0月时与鼠密度相关性最高(表 2)。结合模型的最终拟合效果,选择滞后0月为月平均日照时数的最佳滞后期,其余气象因素均选择滞后2月为最佳滞后期纳入模型。

|
GAM拟合总鼠密度结果显示,滞后2月的累计降水量对鄂尔多斯市总鼠密度的影响无统计学意义,滞后2月的月平均气温、月平均气压与总鼠密度呈非线性关系,月平均日照时数与当月总鼠密度呈线性关系。见图 3。

|
| 注:RR相对危险度;CI置信区间。 图 3 月平均气温、月平均气压和月平均日照时数与总鼠密度的关系 Figure 3 Relationship between monthly average temperature, monthly average air pressure, and monthly average sunshine duration and rodent density |
| |
子午沙鼠与其他鼠种密度的亚组结果显示,滞后2月的月平均气温(F=5.248,P < 0.001)、月平均气压(F=5.281,P=0.001)与当月平均日照时数(F=7.181,P=0.011)对子午沙鼠密度有影响。由图 4可见,随着月平均气温的升高,子午沙鼠密度总体呈下降趋势,在约12 ℃时存在1个小高峰,在18 ℃左右到达最低值,随后又快速上升。子午沙鼠密度随月平均气压先升高后降低,约864 hPa时达到峰值。当月平均日照时数对子午沙鼠密度的影响呈线性关系,随着月平均日照时数的升高,鼠密度逐渐上升。但以上3种因素对其他鼠种密度的影响均无统计学意义(均P > 0.05)。

|
| 注:RR相对危险度;CI置信区间。 图 4 月平均气温、月平均气压和月平均日照时数与子午沙鼠密度(A、B、C)和其他鼠种密度(D、E、F)的关系 Figure 4 Relationship between monthly average temperature, monthly average air pressure, and monthly average sunshine duration and the density of Meriones meridianus (A, B, C) and other rodent species (D, E, F) |
| |
滞后2月的月平均气温和同期的月平均降水量、滞后2月的月平均气温和当月月平均日照时数均对子午沙鼠密度的影响存在交互作用,且有统计学意义(均P < 0.05)(图 5)。在气温约为12 ℃且强降水时,子午沙鼠密度存在1个小高峰;而弱降水条件下,温度对鼠密度的影响较小。当月长平均月日照时数会使得滞后2月的高温和低温条件下的鼠密度均增高,而滞后2月的月平均气温约为12 ℃时,子午沙鼠密度达到短月平均日照时数期间的峰值。

|
| 注:TEM_Avg_lag2表示滞后2月的平均气温;PRE_Time_2020_lag2表示滞后2月时的降水量;SSH表示当月光照时数。 图 5 月平均气温与月累计降水量(A)、月平均日照时数(B)对子午沙鼠密度的交互作用 Figure 5 The interactive effect of monthly cumulative rainfall (A) and monthly average sunshine duration (B) with monthly average temperature on the rodent density |
| |
本研究利用鄂尔多斯市鼠密度和同期气象资料,采用GAM分析了气象因素对鼠密度的直接影响,在此基础上进一步探讨了气象因素间的交互作用对子午沙鼠密度的影响。结果显示,2013-2021年鄂尔多斯市鼠密度整体呈现逐年下降趋势,且鼠密度与气象因素密切相关,滞后2月平均气温与同期累计降水量和当月日照时数对子午沙鼠密度均有一定的交互作用。
本研究结果表明,在2013-2021年鄂尔多斯市总鼠密度逐渐下降,这种趋势可能受到气候变化和人类活动的影响[18]。气候变化可改变鼠类的适生区,进而降低鼠密度[19]。同时,近年来的常态化牧区管理和灭鼠措施等人类活动也可能对鼠类种群密度产生不利影响。本研究发现,2018年鼠密度存在一个峰值,推测这一现象与种群密度的周期性有关[20]。本研究的监测点均位于长爪沙鼠鼠疫自然疫源地,但通过笼捕法得到的优势鼠种为子午沙鼠。另有研究使用5 m夹线法对长爪沙鼠自然疫源地的夜行鼠进行数量监测时,主要捕获到的鼠种也是子午沙鼠[21]。这说明捕获方式和时间对鼠密度监测存在影响,提示在未来的监测过程中,应结合特定地区的实际情况和鼠类的生态习性,选择合适的捕获方式进行鼠密度监测,以确保监测结果更为准确和科学。
本研究发现,滞后2月月平均气温对总鼠密度呈非线性影响,变化趋势与子午沙鼠基本一致。在约-8 ℃时,总鼠密度达到最高峰,此后随着气温的升高,鼠密度呈波动下降趋势,约18 ℃时鼠密度最低,之后迅速升高。而子午沙鼠密度变化差异在于其不存在约-8 ℃的峰值,而在12 ℃左右存在1个较高的峰值,这可能是与鼠类的季节性变化和生境适宜性有关。冬季温度上升可能干扰了其他鼠类的冬眠状态[22],从而限制了子午沙鼠这类非冬眠鼠种的生存,进而导致春季其密度的下降。春季对夏季鼠密度的影响呈先升后降的趋势,约在12 ℃时达到最高点。春季温度升高引发鼠类繁殖,种群密度增加,但密度上升导致资源竞争激化,达到临界点后竞争限制种群增长,进而导致下降。Eccard等[23]研究发现春季的种群竞争导致雄性田鼠的数量显著减少。夏季温度约从18 ℃开始逐渐升高会导致秋季鼠密度快速增加。温度升高促进植物生长和繁殖,提供更丰富的食物资源,这使得鼠类在较高温度下更易获取充足的食物,获得能量和营养,支持其生存和繁殖,进而导致秋季鼠密度的迅速增加。郭强等[24]对高原鼢鼠(Eospalax baileyi)的研究发现,温度主要通过对食物的影响进而对其种群密度产生间接影响。
目前关于气压对鼠类的影响研究较少,主要集中在气压对鼠类肠道微生物的研究,提示鼠类某些胃肠道疾病可能与气压过低或者过高有关[25-26]。本研究结果显示,随着滞后2月的月平均气压逐渐升高,子午沙鼠密度先升高后下降,大约在864 hPa达到峰值。这与王金娜等[10]的研究基本一致,月平均水汽压对鼠密度有滞后效应,即在前1个月的月平均水汽压升高的情况下,鼠密度也会在接下来的1个月内上升。鼠类可能对低气压升高更为敏感,气压升高会刺激它们的生态活动,导致鼠密度的逐渐升高。然而,随着气压继续升高,其他不利因素可能开始发挥作用,抵消了气压对鼠密度的正向影响,从而导致鼠密度开始下降。需要注意的是,前述解释是基于统计模型的推测,鼠类对气压的生态响应涉及众多复杂的生物学机制,需要进一步的研究来深入探讨。
本研究发现随着月平均日照时数的延长,鼠密度呈线性增长。从鼠类自身的生理出发,有研究者发现,长日照在小鼠维生素D的合成中起重要作用[27]。Gaston和Menaker[28]研究发现,成年仓鼠的睾丸每天至少需要12.5 h的光照才能维持精子形成并防止退化。而从生态学角度来看,这一现象可借助营养级联反应的理论来解释[29],即随着日照时数的延长,植物光合作用的时间增加,导致植物生长和繁殖的增加,这使得鼠类的食物资源变得更加丰富和可获得,从而支持鼠类的生存和繁殖。
气象因素对鼠密度有着复杂的作用机制,本研究结果显示滞后2月的月平均气温与其同期累计降水量和当月月平均日照时数对鼠密度存在一定交互作用。月累计降水量的单独作用无统计学意义,但与月平均气温的交互作用分析表明,适宜温度的强降水量使得鼠密度增高。这可能是啮齿动物通常受益于降水的增加,特别是在典型的干燥生境,高降水的“好年景”改善了它们的食草和水平衡[30]。长日照时数下的低温和高温都可能导致鼠密度上升,可能因其在这种条件下准备越冬和越冬后觅食的活动增加。此外,适宜温度下的短月平均日照时数导致鼠密度增加,可能是春季适宜温度促进了子午沙鼠生长发育,而当在夏季遇短日照情况时,其夜行习性使其在夜间更活跃[31]。
综上所述,月平均气温和月平均气压对鄂尔多斯市鼠密度呈非线性影响,而月平均日照时数对鼠密度呈线性影响,且月平均气温、月累计降水量和月平均日照时数之间对鼠密度存在一定的交互作用。本研究结果可为鼠密度监测研究提供一定的参考,并提示在今后的防鼠灭鼠工作中,应充分利用鼠密度监测数据,结合相关气象条件综合考虑,通过多项举措减轻鼠类危害,从而有效预防和控制鼠源疾病。
利益冲突 无
| [1] |
Sun Z, Xu L, Schmid BV, et al. Human plague system associated with rodent diversity and other environmental factors[J]. Roy Soc Open Sci, 2019, 6(6): 190216. DOI:10.1098/rsos.190216 |
| [2] |
Xiao H, Tong X, Gao LD, et al. Spatial heterogeneity of hemorrhagic fever with renal syndrome is driven by environmental factors and rodent community composition[J]. PLoS Negl Trop Dis, 2018, 12(10): e0006881. DOI:10.1371/journal.pntd.0006881 |
| [3] |
Vitale M, Agnello S, Chetta M, et al. Human leptospirosis cases in Palermo Italy. The role of rodents and climate[J]. J Infect Public Health, 2018, 11(2): 209-214. DOI:10.1016/j.jiph.2017.07.024 |
| [4] |
刘起勇. 气候变化对中国媒介生物传染病的影响及应对: 重大研究发现及未来研究建议[J]. 中国媒介生物学及控制杂志, 2021, 32(1): 1-11. Liu QY. Impact of climate change on vector-borne diseases and related response strategies in China: Major research findings and recommendations for future research[J]. Chin J Vect Biol Control, 2021, 32(1): 1-11. DOI:10.11853/j.issn.1003.8280.2021.01.001 |
| [5] |
段天一, 宋驰, 赵玮, 等. 2003-2021年人间鼠疫发病月份及相关因素对比分析[J]. 中国地方病防治, 2022, 37(6): 467-469. Duan TY, Song C, Zhao W, et al. Analysis of the incidence months of human plague from 2003 to 2021[J]. Chin J Control Endem Dis, 2022, 37(6): 467-469. |
| [6] |
方喜业, 杨瑞馥, 许磊, 等. 中国鼠疫自然疫源地分型研究Ⅶ. 中国鼠疫自然疫源地分型生物学特征[J]. 中华流行病学杂志, 2012, 33(11): 1144-1150. Fang XY, Yang RF, Xu L, et al. Ecological-geographic landscapes of natural plague foci in China Ⅶ. Typing of natural plague foci[J]. Chin J Epidemiol, 2012, 33(11): 1144-1150. DOI:10.3760/cma.j.issn.0254-6450.2012.11.011 |
| [7] |
牛艳芬, 张懿晖, 康东梅, 等. 河北省长爪沙鼠及其巢蚤密度与气象因素关系[J]. 中国媒介生物学及控制杂志, 2022, 33(5): 684-688. Niu YF, Zhang YH, Kang DM, et al. Relationship between population densities of Meriones unguiculatus and its nest fleas and meteorological factors in M. unguiculatus plague foci of Hebei Province, China[J]. Chin J Vect Biol Control, 2022, 33(5): 684-688. DOI:10.11853/j.issn.1003.8280.2022.05.013 |
| [8] |
刘自远. 气象因素对农田鼠类数量影响的典型相关分析[J]. 中国媒介生物学及控制杂志, 2011, 22(4): 372-374. Liu ZY. Canonical correlation analysis of climate factors and farmland rodent density[J]. Chin J Vect Biol Control, 2011, 22(4): 372-374. |
| [9] |
Jiang GS, Liu J, Xu L, et al. Climate warming increases biodiversity of small rodents by favoring rare or less abundant species in a grassland ecosystem[J]. Integr Zool, 2013, 8(2): 162-174. DOI:10.1111/1749-4877.12027 |
| [10] |
王金娜, 劳家辉, 侯娟, 等. 气象因素对鼠密度的影响研究[J]. 预防医学, 2018, 30(9): 870-873, 878. Wang JN, Lao JH, Hou J, et al. Effect of meteorological factors on rodent density[J]. Prev Med, 2018, 30(9): 870-873, 878. DOI:10.19485/j.cnki.issn2096-5087.2018.09.002 |
| [11] |
武晓东, 袁帅, 付和平, 等. 不同干扰下阿拉善荒漠啮齿动物优势种对气候变化的响应[J]. 生态学报, 2016, 36(6): 1765-1773. Wu XD, Yuan S, Fu HP, et al. Responses of dominant rodent species to climate change in different disturbed habitats in the Alashan desert[J]. Acta Ecol Sin, 2016, 36(6): 1765-1773. DOI:10.5846/stxb201408061564 |
| [12] |
Liu QY, Xu WB, Lu S, et al. Landscape of emerging and re-emerging infectious diseases in China: Impact of ecology, climate, and behavior[J]. Front Med, 2018, 12(1): 3-22. DOI:10.1007/s11684-017-0605-9 |
| [13] |
侯祥, 韩宁, 封托, 等. 气候因素对黑线姬鼠种群动态影响的非线性效应[J]. 生态学报, 2020, 40(14): 4836-4841. Hou X, Han N, Feng T, et al. Nonlinear effects of climate on population dynamics of Apodemus agrarius[J]. Acta Ecol Sin, 2020, 40(14): 4836-4841. DOI:10.5846/stxb201904240836 |
| [14] |
吴海磊, 钱吉生, 阮治安, 等. 鼠密度与气象因素的响应面分析研究[J]. 中国媒介生物学及控制杂志, 2009, 20(2): 129-132. Wu HL, Qian JS, Ruan ZA, et al. Study on the relationship of meteorological factors and rats density by response surface methodology[J]. Chin J Vect Biol Control, 2009, 20(2): 129-132. |
| [15] |
Previtali MA, Lima M, Meserve PL, et al. Population dynamics of two sympatric rodents in a variable environment: Rainfall, resource availability, and predation[J]. Ecology, 2009, 90(7): 1996-2006. DOI:10.1890/08-0405.1 |
| [16] |
Wan XR, Yan C, Wang ZY, et al. Sustained population decline of rodents is linked to accelerated climate warming and human disturbance[J]. BMC Ecol Evol, 2022, 22(1): 102. DOI:10.1186/s12862-022-02056-z |
| [17] |
Ver Hoef JM, Boveng PL. Quasi-Poisson vs. negative binomial regression: How should we model overdispersed count data?[J]. Ecology, 2007, 88(11): 2766-2772. DOI:10.1890/07-0043.1 |
| [18] |
李紫薇, 尹家祥. 鼠类与环境因素和人类活动的关系[J]. 中国媒介生物学及控制杂志, 2023, 34(1): 117-121. Li ZW, Yin JX. The relationship between rodents and environmental factors and human activities[J]. Chin J Vect Biol Control, 2023, 34(1): 117-121. DOI:10.11853/j.issn.1003.8280.2023.01.022 |
| [19] |
Xu L, Wang Q, Yang RF, et al. Climate-driven marmot-plague dynamics in Mongolia and China[J]. Sci Rep, 2023, 13(1): 11906. DOI:10.1038/s41598-023-38966-1 |
| [20] |
Andreassen HP, Sundell J, Ecke F, et al. Population cycles and outbreaks of small rodents: Ten essential questions we still need to solve[J]. Oecologia, 2021, 195(3): 601-622. DOI:10.1007/s00442-020-04810-w |
| [21] |
韩冰, 刘慧杰, 张大禹, 等. 内蒙古自治区2018-2022年长爪沙鼠鼠疫疫源地动物间鼠疫流行病学调查[J]. 中国媒介生物学及控制杂志, 2023, 34(5): 697-702. Han B, Liu HJ, Zhang DY, et al. An epidemiological survey of animal plague in Meriones unguiculatus plague foci of Inner Mongolia Autonomous Region, China, 2018-2022[J]. Chin J Vect Biol Control, 2023, 34(5): 697-702. DOI:10.11853/j.issn.1003.8280.2023.05.020 |
| [22] |
MacCannell ADV, Staples JF. Elevated ambient temperature accelerates aspects of torpor phenology in an obligate hibernator[J]. J Therm Biol, 2021, 96: 102839. DOI:10.1016/j.jtherbio.2021.102839 |
| [23] |
Eccard JA, Fey K, Caspers BA, et al. Breeding state and season affect interspecific interaction types: Indirect resource competition and direct interference[J]. Oecologia, 2011, 167(3): 623-633. DOI:10.1007/s00442-011-2008-y |
| [24] |
郭强, 王玉琴, 鲍根生, 等. 气象因子对高原鼢鼠种群数量的影响[J]. 草业学报, 2020, 29(8): 188-194. Guo Q, Wang YQ, Bao GS, et al. Effect of meteorological factors on the population of plateau zokor[J]. Acta Pratacul Sin, 2020, 29(8): 188-194. DOI:10.11686/cyxb2019474 |
| [25] |
Maity C, Adak A, Pathak TK, et al. Study of the cultivable microflora of the large intestine of the rat under varied environmental hyperbaric pressures[J]. J Microbiol Immunol Infect, 2012, 45(4): 281-286. DOI:10.1016/j.jmii.2011.12.002 |
| [26] |
Adak A, Maity C, Ghosh K, et al. Alteration of predominant gastrointestinal flora and oxidative damage of large intestine under simulated hypobaric hypoxia[J]. Z Gastroenterol, 2014, 52(2): 180-186. DOI:10.1055/s-0033-1336007 |
| [27] |
Lin MY, Lim LM, Tsai SP, et al. Low dose ultraviolet B irradiation at 308 nm with light-emitting diode device effectively increases serum levels of 25(OH)D[J]. Sci Rep, 2021, 11(1): 2583. DOI:10.1038/s41598-021-82216-1 |
| [28] |
Gaston S, Menaker M. Photoperiodic control of hamster testis[J]. Science, 1967, 158(3803): 925-928. DOI:10.1126/science.158.3803.925 |
| [29] |
Xu L, Schmid BV, Liu J, et al. The trophic responses of two different rodent-vector-plague systems to climate change[J]. Proc Biol Sci: Biol Sci, 2015, 282(1800): 20141846. DOI:10.1098/rspb.2014.1846 |
| [30] |
Biggins DE, Eads DA, Godbey JL. Plague transforms positive effects of precipitation on prairie dogs to negative effects[J]. Int J Parasitol Parasites Wildl, 2021, 14: 329-334. DOI:10.1016/j.ijppaw.2021.02.0 |
| [31] |
Yang SW, Yuan S, Wu XD, et al. The effect of grazing on winter survival of midday gerbil (Meriones meridianus) of different genders[J]. Ecol Evol, 2020, 10(21): 12395-12406. DOI:10.1002/ece3.6870 |