 2023, Vol. 34
2023, Vol. 34扩展功能
文章信息
- 王玲, 何亚明, 季恒青, 李志峰, 赵婷, 黄为, 肖汉森
- WANG Ling, HE Ya-ming, JI Heng-qing, LI Zhi-feng, ZHAO Ting, HUANG Wei, XIAO Han-sen
- 三峡库区重庆段2019-2021年小型兽类及体表媒介蚤监测结果分析
- An analysis of surveillance results of small mammals and body surface fleas in the Chongqing section of the Three Gorges Reservoir Region, China, 2019-2021
- 中国媒介生物学及控制杂志, 2023, 34(5): 642-647
- Chin J Vector Biol & Control, 2023, 34(5): 642-647
- 10.11853/j.issn.1003.8280.2023.05.011
-
文章历史
- 收稿日期: 2023-04-11



2 高致病性病原微生物重庆市重点实验室, 重庆 400042;
3 重庆市疾病预防与公共卫生研究中心, 重庆 400042;
4 重庆市疾病预防控制中心 消毒与媒介生物控制所, 重庆 400042
2 Chongqing Municipal Key Laboratory for High Pathogenic Microbes, Chongqing 400042, China;
3 Chongqing Center for Disease Prevention and Public Health Research, Chongqing 400042, China;
4 Department of Disinfection and Vector Control, Chongqing Center for Disease Control and Prevention, Chongqing 400042, China
三峡库区是指三峡水利枢纽工程中三峡大坝蓄水达175 m后,因水位升高而被淹受影响的有关行政区域。三峡库区是中国乃至全世界最为特殊的生态功能区,其水土保持和生态稳定对三峡水利工程的长期安全运行及长江中下游的生态安全有重大战略意义。三峡库区重庆段覆盖了整个三峡库区总面积的85.6%,占据了极其重要的生态地理位置[1]。三峡大坝水库蓄水形成后,生态环境发生了一定改变,水位从2008年起正常蓄水达到175 m,且三峡水库实行“冬储夏排”的运行模式,水位改变对于栖息在库区的小型兽类(小兽)的生存空间造成影响。以鼠类为主的小兽及其体表媒介蚤有可能会引起鼠疫、肾综合征出血热、钩端螺旋体病、地方性斑疹伤寒和隐孢子虫病等多种鼠传人兽共患病的发生流行[2]。我国从2004年开始开展三峡库区鼠疫防治监测项目,对该地区的小兽种类构成、密度及重要鼠源疾病发病情况进行了持续监测,以期对鼠疫等相关疾病提供基础资料和科学指导。现将2019-2021年三峡库区重庆段小兽及体表媒介蚤的监测结果分析如下。
1 材料与方法 1.1 资料来源数据来源于2019-2021年重庆市三峡库区重庆段小兽及体表媒介蚤监测点监测数据。
1.2 监测点概况按照《重庆市鼠传疾病防治项目监测方案》要求,在三峡库区重庆段选择8个县(区)(涪陵、丰都、忠县、万州、云阳、开州、奉节和巫山),每个县(区)各选择2~3个监测点开展监测,监测点包括居住区和耕作地2类生境,距水库淹没线1 km以内。于每年1、3、5、7、9、11月各开展1次小兽种群密度监测和体表寄生蚤监测。
1.3 监测方法 1.3.1 小兽监测采用夹(笼)夜法,每个监测点分生境各布放100个鼠夹(笼),诱饵使用卤猪头肉(皮面积1 cm×1 cm),晚放晨收,连续监测3 d。将捕获小兽进行分类,计算密度。
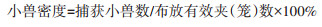
|
将捕获小兽单只装入布袋,用乙醚麻醉,梳检体表寄生蚤,置于蚤管内待检,计算染蚤率及蚤指数。

|
各监测点对捕获的小兽及周边地区鼠疫易感动物(与鼠类接触有可能被感染的犬、猫及其他动物)抽血,采用鼠疫间接血凝试验检测鼠疫F1抗体。
1.4 统计学分析将监测资料归纳整理,形成Excel分析用数据库。利用SPSS 25.0软件进行统计分析,小兽密度、构成比、染蚤率等率的比较采用χ2检验,P < 0.05为差异有统计学意义。
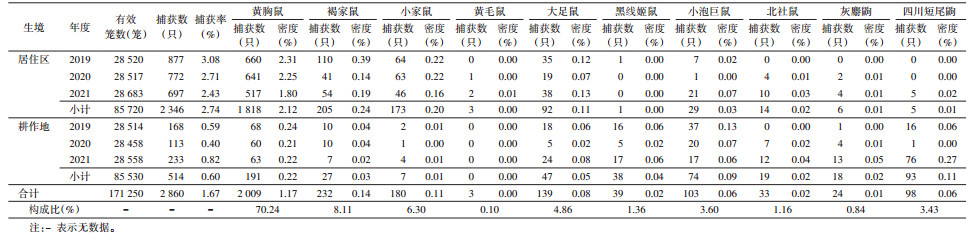
2 结果 2.1 小兽构成共布放有效鼠夹(笼)171 250个,捕获小兽2 860只,平均密度为1.67%。在居住区和耕作地2种生境共发现小兽10种,其中9种为二者共有(表 1)。各小兽的构成比依次为黄胸鼠(Rattus tanezumi)70.24%、褐家鼠(R. norvegicus)8.11%、小家鼠(Mus musculus)6.30%、大足鼠(R. nitidus)4.86%、小泡巨鼠(Leopoldamys edwardsi)3.60%、四川短尾鼩(Anourosorex squamipes)3.43%、黑线姬鼠(Apodemus agrarius)1.36%、北社鼠(Niviventer confucianus)1.16%、灰麝鼩(Crocidura attenuata)0.84%和黄毛鼠(R. losea)0.10%。
|
居住区共布放有效鼠夹(笼)85 720个,捕获小兽2 346只,平均小兽密度为2.74%;2019-2021年各年居住区小兽密度有差异,呈下降趋势,差异有统计学意义(χ2=22.493,P < 0.001);小兽种类以黄胸鼠(77.49%)、褐家鼠(8.74%)和小家鼠(7.37%)为主。在耕作地布放有效鼠夹(笼)85 530个,捕获小兽514只,平均小兽密度为0.60%;2019-2021年耕作地小兽密度在较低水平波动,各年小兽密度差异有统计学意义(χ2=41.953,P < 0.001);小兽以黄胸鼠(37.16%)、四川短尾鼩(18.09%)、小泡巨鼠(14.40%)为主。居住区与耕作地小兽构成比差异有统计学意义(χ2=1 003.754,P < 0.001)。
2.3 三峡库区重庆段不同地区小兽构成及密度监测的8个县(区)中,居住区捕获小兽种类最多的是涪陵区(9种),各县(区)居住区鼠密度最高的均为黄胸鼠;耕作地捕获小兽种类最多的是万州区(8种)。见图 1、2。

|
| 图 1 2019-2021年三峡库区重庆段不同地区居住区小型兽类种类构成及密度 Figure 1 Species composition and density of small mammals in residential areas of different places of the Chongqing section of the Three Gorges Reservoir Region, 2019-2021 |
| |

|
| 图 2 2019-2021年三峡库区重庆段不同地区耕作地小型兽类种类构成及密度 Figure 2 Species composition and density of small mammals in cropland areas of different places of the Chongqing section of the Three Gorges Reservoir Region, 2019-2021 |
| |
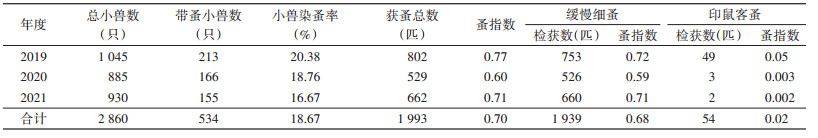
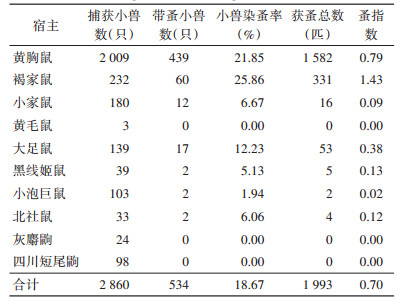
共梳检小兽2 860只,带蚤小兽534只,共获蚤1 993匹,小兽染蚤率为18.67%,总蚤指数0.70;年染蚤率2019-2021年从20.38%降低至16.67%,差异无统计学意义(χ2=4.481,P=0.106),蚤指数在0.60~0.77波动;检获蚤种有缓慢细蚤(Leptothris segnis)和印鼠客蚤(Xenopsylla cheopis)2种,蚤指数分别为0.68和0.02(表 2)。不同小兽中褐家鼠染蚤率最高,为25.86%,蚤指数为1.43,黄胸鼠次之,染蚤率为21.85%,蚤指数0.79(表 3)。鼠体染蚤率和蚤指数变化具有季节性,均以3月最高,染蚤率为29.05%,蚤指数为1.35;其次是1月,染蚤率为24.69%,蚤指数为0.83。见图 3。
|
|

|
| 图 3 2019-2021年三峡库区重庆段不同月份小型兽类染蚤率及蚤指数 Figure 3 Flea infestation rates and flea index values of small mammals by month in the Chongqing section of the Three Gorges Reservoir Region, 2019-2021 |
| |
各监测点捕获小兽2 860只,采获易感动物(犬、猫)262只,均抽血进行了鼠疫间接血凝试验检测F1抗体,结果均为阴性。
3 讨论长江三峡水利枢纽工程于1994年正式动工,2003年首次蓄水,2009年全部竣工并正式运行。三峡库区重庆段位于北纬28°31'~31°44',东经105°49'~110°12',其动物地理区划属东洋界华中区西部山地高原亚区,与广西壮族自治区和贵州省相同,具有南方家鼠鼠疫疫源地类似的地理条件[3-4]。三峡水库的形成给库区带来一系列气候、环境及生态变化,可能导致某些传染病的发生流行[5]。
毛德强等[6] 1997-2012年鼠疫监测报道,成库初期三峡库区重庆段居住区的优势鼠种为褐家鼠、小家鼠,耕作地的优势鼠种为四川短尾鼩、褐家鼠,且成库后鼠密度保持在较低水平。2019-2021年的监测结果显示,三峡库区重庆段小兽在居住区以黄胸鼠、褐家鼠为主,耕作地以黄胸鼠、四川短尾鼩为主,优势鼠种与成库初期相比有较明显的变化。居住区鼠密度呈现下降趋势,耕作地鼠密度保持在较低水平波动,这与三峡库区近年来开展的鼠疫监测结果相符合[7-8]。主要原因为三峡工程完成常规蓄水及运行后,每年库区两岸周而复始水位的升高降落形成水陆交替的消落带,随着消落带水沙、植被、土壤、微生物等生态改变[9-11],鼠类发生迁移,影响了小兽种群构成及密度。
2019-2021年监测发现,三峡库区重庆段居住区和耕作地2种生境的优势鼠种均为黄胸鼠。黄胸鼠是我国主要家栖鼠种之一,分布属东南亚热带-亚热带型,在我国主要分布于长江流域以南地区,随着城市化进程中环境的变迁、气候变暖、交通便利以及人为因素的影响,20世纪80、90年代在我国陕西、山西、甘肃、山东省、宁夏回族自治区和新疆维吾尔自治区等地均有报道,该鼠种明显表现出北扩趋势,在南方部分地区,黄胸鼠的优势地位已被其他鼠种所取代[12]。但三峡库区自2011年以来的监测结果表明黄胸鼠的密度、构成均明显上升[6-7]。黄胸鼠密度及构成比大幅上升,原因可能与以下几方面有关: 一是捕鼠效率提高,捕鼠方法从鼠夹法(2010年以前)变为鼠笼法[13]。二是三峡工程施工后于1998-2008年间开展了4次库底卫生清理及大规模灭鼠工作,迅速降低了库区鼠密度,2009年长江三峡水利工程竣工后未再对库区采取大规模灭鼠措施,鼠类繁殖生长旺盛。三是三峡移民搬迁房屋重建,所建住房大多是砖混结构的楼房,屋顶一般构建木质大梁,其上覆盖瓦片,房屋顶端形成阁楼或夹层,满足黄胸鼠生活习性,有利于黄胸鼠生存繁殖;同时就地安置移民,人群居住集中,人居密度增加,更有利于黄胸鼠获取食物。黄胸鼠虽作为家栖鼠种,但仍然具有一定的野栖性,监测显示,黄胸鼠已成为当前耕作地的优势鼠种,家鼠野鼠种间接触频繁,鼠疫等自然疫源性疾病传播风险增大。
三峡库区重庆段媒介蚤监测发现有缓慢细蚤、印鼠客蚤,优势蚤种为缓慢细蚤,印鼠客蚤构成及蚤指数均较低。何晋侯等[14]通过实验发现印鼠客蚤菌栓形成率较高,是黄胸鼠鼠疫的主要媒介蚤;缓慢细蚤媒介效能极低,为家鼠鼠疫的次要媒介。但蚤的媒介效能与宿主种类、鼠疫菌株类型以及疫源地的生态环境等因素有关[15]。监测发现,蚤指数变化具有季节性,与国内有些地区蚤密度夏秋较高,冬春较低有所不同[16],库区重庆段蚤指数3月最高,蚤指数为1.35,其次是1月,蚤指数为0.83。有研究表明,气温、环境湿度影响蚤类的生存发育且不同季节蚤类繁殖力不同,蚤类群落因此会发生季节性变化[17-19]。重庆市位于长江峡谷,湿度高,入夏早,高温持续时间长,这些特殊的气候条件可能导致鼠蚤的繁殖发育不同于国内其他地区[18, 20]。
三峡库区重庆段具有南方家鼠鼠疫自然疫源地存在的相应自然地理景观,黄胸鼠广泛分布于监测区域内,参照大型建设项目鼠疫卫生学评价风险评估标准[21],黄胸鼠室内密度为2.12%,已远超国家控制标准(1.0%);黄胸鼠蚤指数为0.79,临近鼠疫警戒指标(1)。鼠疫的传播和流行以及鼠疫菌的持续存在与当地生态变化、生态稳定性以及宿主和媒介蚤类多样性相关[22-23]。2019-2021年三峡库区重庆段小兽种群及密度与成库时相比均发生了明显变化,宿主动物血清学检测鼠疫F1抗体虽未发现阳性,但主要宿主黄胸鼠密度明显高于国家相关控制标准和主要媒介蚤类印鼠客蚤的存在,提示该区域为潜在危险地区,应引起高度关注。鼠疫的传播方式是“宿主动物-传播媒介-宿主动物”,且鼠疫的发生具有突发性,在鼠疫自然疫源地常表现为流行期短而间歇期长的特征,鼠疫发生后常在追溯调查后才得以证实,对动物及人类存在巨大的潜在危害,因此鼠疫的预警和早期干预尤为重要。在三峡库区重庆段开展动物鼠疫监测的同时需开展灭鼠灭蚤活动,以期降低鼠密度,有效遏制鼠患,进而降低鼠疫发生风险。
鼠疫为自然疫源性疾病,鼠疫的发生和流行与气候条件、地理环境、生态环境等息息相关。三峡水库的蓄水及“冬储夏排”的运行模式,在库区沿岸形成周而复始的消落区,水位的上升降落,对库区周边生态、地质及气候等产生了直接影响,小兽种群分布及密度均发生了较大变化,体表媒介蚤也跟随宿主的变化而发生动态改变。有文献报道,目前在三峡库区的研究热点主要集中在植物群落演变过程和土壤环境变化,而对动物、微生物的生态演变研究较少[24]。因此,应该在三峡库区长期开展小兽及其体表媒介蚤的监测,密切关注其变化趋势,可在鼠疫等自然疫源性疾病发生的早期起到预警及干预作用,同时也能为新发及再发其他鼠源疾病的诊断、溯源和干预提供基础资料和本底数据。
志谢 重庆市鼠传疾病防治监测项目得到各县(区)疾病预防控制中心大力支持,衷心感谢利益冲突 无
| [1] |
王丽. 基于生态足迹模型的重庆三峡库区的可持续发展研究[D]. 重庆: 西南大学, 2012. Wang L. Analysis of sustainable development of TGP reservoir area based on ecological footprint model[D]. Chongqing: Southwest University, 2012. (in Chinese) |
| [2] |
徐爱玲, 栗冬梅, 刘起勇. "鼠源疾病"的概念与内涵的系统评价[J]. 中国媒介生物学及控制杂志, 2022, 33(2): 161-170. Xu AL, Li DM, Liu QY. The concept and connotation of "rodent-borne diseases": A systematic study[J]. Chin J Vector Biol Control, 2022, 33(2): 161-170. DOI:10.11853/j.issn.1003.8280.2022.02.001 |
| [3] |
宋开莲, 贺启生, 吴定昌, 等. 2006-2015年贵州省黔西南州鼠疫监测结果分析[J]. 中华地方病学杂志, 2018, 37(6): 493-496. Song KL, He QS, Wu DC, et al. Analysis of the plague surveillance results in Qianxi'nan Prefecture, Guizhou province from 2006 to 2015[J]. Chin J Endemiol, 2018, 37(6): 493-496. DOI:10.3760/cma.j.issn.2095-4255.2018.06.014 |
| [4] |
陆献蒿, 余水兰, 周建松. 2008-2014年广西百色市鼠疫监测分析[J]. 寄生虫病与感染性疾病, 2016, 14(1): 33-36. Lu XH, Yu SL, Zhou JS. Plague surveillance in Baise, Guangxi, 2008-2014[J]. Parasit Infect Dis, 2016, 14(1): 33-36. |
| [5] |
Guo YH, Lai SJ, Zhang J, et al. Mosquito population dynamics during the construction of Three Gorges Dam in Yangtze River, China[J]. Acta Trop, 2018, 182: 251-256. DOI:10.1016/j.actatropica.2018.03.008 |
| [6] |
毛德强, 李洪, 张春华, 等. 三峡库区成库前后鼠疫相关鼠形动物种群及数量变化趋势[J]. 中国媒介生物学及控制杂志, 2016, 27(1): 68-70. Mao DQ, Li H, Zhang CH, et al. Rodent charateristic changes in the Three Gorges Reservoir before and after the water storage[J]. Chin J Vector Biol Control, 2016, 27(1): 68-70. DOI:10.11853/j.issn.1003.4692.2016.01.022 |
| [7] |
何亚明, 毛德强, 季恒青, 等. 三峡库区重庆段鼠形动物种群分布及密度变化研究[J]. 中国媒介生物学及控制杂志, 2020, 31(5): 580-586. He YM, Mao DQ, Ji HQ, et al. A study of the population distribution and density changes of mouse-like rodents in the Chongqing section of the Three Gorges Reservoir area, China[J]. Chin J Vector Biol Control, 2020, 31(5): 580-586. DOI:10.11853/j.issn.1003.8280.2020.05.016 |
| [8] |
赵鑫, 刘建华, 张皓. 三峡库区宜昌段1997-2020年鼠形动物监测报告[J]. 中国媒介生物学及控制杂志, 2022, 33(1): 67-71. Zhao X, Liu JH, Zhang H. Surveillance report of rodents in Yichang section of the Three Gorges Reservoir area, 1997-2020[J]. Chin J Vector Biol Control, 2022, 33(1): 67-71. DOI:10.11853/j.issn.1003.8280.2022.01.012 |
| [9] |
林莉, 董磊, 潘雄, 等. 三峡水库蓄水后库区水沙变化及其生态环境响应特征[J]. 湖泊科学, 2023, 35(2): 411-422. Lin L, Dong L, Pan X, et al. Water and sediment changes and the eco-environment response characteristics in Three Gorges Reservoir after the impoundment[J]. J Lake Sci, 2023, 35(2): 411-422. DOI:10.18307/2023.0204 |
| [10] |
程莅登. 长江重庆段河岸植物群落及物种多样性研究[D]. 重庆: 西南大学, 2019. Cheng LD. Study on riparian plant community and species diversity in Chongqing section of the Yangtze River[D]. Chongqing: Southwest University, 2019. (in Chinese) |
| [11] |
叶飞. 三峡库区消落带土壤氮循环关键过程微生物群落特征研究[D]. 北京: 中国科学院大学(中国科学院重庆绿色智能技术研究院), 2018. Ye F. Microbial community characteristics of the key processes in soil nitrogen cycle in the water-level fluctuation zone of the Three Gorges Reservoir area[D]. Beijing: University of Chinese Academy of Sciences (Chongqing Institute of Green and Intelligent Technology, Chinese Academy of Sciences), 2018. (in Chinese) |
| [12] |
张美文, 郭聪, 王勇, 等. 我国黄胸鼠的研究现状[J]. 动物学研究, 2000, 21(6): 487-497. Zhang MW, Guo C, Wang Y, et al. The buff-breasted rats (Rattus flavipectus) in China[J]. Zool Res, 2000, 21(6): 487-497. DOI:10.3321/j.issn:0254-5853.2000.06.013 |
| [13] |
章士军, 王智泉, 陈郁, 等. 南通市区鼠夹法和鼠笼法捕鼠情况的分析[J]. 中华卫生杀虫药械, 2013, 19(2): 153-154. Zhang SJ, Wang ZQ, Chen Y, et al. Mouse density survey with snap trap and cage trap in Nantong city[J]. Chin J Hyg Insect Equip, 2013, 19(2): 153-154. DOI:10.19821/j.1671-2781.2013.02.021 |
| [14] |
何晋侯, 梁云, 张洪英, 等. 云南省家野两型疫源地七种主要蚤类传播鼠疫的实验研究[J]. 中华流行病学杂志, 1997, 18(4): 236-240. He JH, Liang Y, Zhang HY, et al. A study on the transmission of plague though seven kinds of fleas in rat type and wild rodent type plague foci in Yunnan[J]. Chin J Epidemiol, 1997, 18(4): 236-240. |
| [15] |
田洁, 任振宁, 鞠成, 等. 蚤类传播鼠疫媒介效能及其影响因素的研究进展[J]. 中国地方病防治, 2022, 37(2): 104-106. Tian J, Ren ZN, Ju C, et al. Research progress on vector efficacy of fleas in plague transmission and its influencing factors[J]. Chin J Ctrl Endem Dis, 2022, 37(2): 104-106. |
| [16] |
康东梅, 闫东, 候芝林, 等. 河北省鼠疫自然疫源地康保牧场长爪沙鼠寄生蚤动态分析[J]. 中国媒介生物学及控制杂志, 2022, 33(1): 72-75. Kang DM, Yan D, Hou ZL, et al. Dynamic analysis of parasitic fleas of Meriones unguiculatus in natural plague focus Kangbao pasture in Hebei province, China[J]. Chin J Vector Biol Control, 2022, 33(1): 72-75. DOI:10.11853/j.issn.1003.8280.2022.01.013 |
| [17] |
杨创明. 四川青海田鼠鼠疫疫源地蚤类群落结构及其动态[D]. 成都: 西南民族大学, 2013. Yang CM. Community structure and dynamics of fleas in Microtus fuscus nature plague foci in Shiqu of China's Sichuan[D]. Chengdu: Southwest University for Nationalities, 2013. (in Chinese) |
| [18] |
高子厚, 刘正祥, 杜春红, 等. 云南居民区鼠类体外寄生蚤物种多样性调查[J]. 昆虫学报, 2014, 57(2): 257-264. Gao ZH, Liu ZX, Du CH, et al. An investigation of species diversity of ectoparasite fleas on rodents in residential areas in Yunnan province, southwestern China[J]. Acta Entomol Sin, 2014, 57(2): 257-264. DOI:10.16380/j.kcxb.2014.02.011 |
| [19] |
Zhang ZB, Li ZQ, Tao Y, et al. Relationship between increase rate of human plague in China and global climate index as revealed by cross-spectral and cross-wavelet analyses[J]. Integr Zool, 2007, 2(3): 144-153. DOI:10.1111/j.1749-4877.2007.00061.x |
| [20] |
白雪薇, 牛艳芬, 陈永明, 等. 1990-2020年河北省鼠疫自然疫源地长爪沙鼠体蚤和巢蚤数量分析[J]. 中华卫生杀虫药械, 2022, 28(3): 213-217. Bai XW, Niu YF, Chen YM, et al. Analyses on the quantity of body fleas and nest fleas of Meriones unguiculatus in the plague natural foci in Hebei province from 1990 to 2020[J]. Chin J Hyg Insect Equip, 2022, 28(3): 213-217. DOI:10.19821/j.1671-2781.2022.03.007 |
| [21] |
卫生部卫生应急办公室, 中国疾病预防控制中心. 鼠疫防控应急手册[M]. 北京: 北京大学医学出版社, 2009: 84-90. Health Emergency Office of the Ministry of Health, Chinese Center for Disease Prevention and Control. Plague prevention and control emergency handbook[M]. Beijing: Peking University Medical Press, 2009: 84-90. |
| [22] |
Eisen RJ, Borchert JN, Mpanga JT, et al. Flea diversity as an element for persistence of plague bacteria in an East African plague focus[J]. PLoS One, 2012, 7(4): e35598. DOI:10.1371/journal.pone.0035598 |
| [23] |
Allen T, Murray KA, Zambrana-Torrelio C, et al. Global hotspots and correlates of emerging zoonotic diseases[J]. Nat Commun, 2017, 8(1): 1124. DOI:10.1038/s41467-017-00923-8 |
| [24] |
王晓锋, 李贤祥, 刘婷婷, 等. 基于CiteSpace的三峡库区消落带研究热点与进展[J]. 生态科学, 2022, 41(1): 249-261. Wang XF, Li XX, Liu TT, et al. Trends and hotspots analysis of the researches on the drawdown zone of Three Gorges Reservoir based on CiteSpace[J]. Ecol Sci, 2022, 41(1): 249-261. DOI:10.14108/j.cnki.1008-8873.2022.01.029 |