 2023, Vol. 34
2023, Vol. 34扩展功能
文章信息
- 谢梦琪, 寸得娇, 姚晓燕, 王飞, 李兰花, 田娜
- XIE Meng-qi, CUN De-jiao, YAO Xiao-yan, WANG Fei, LI Lan-hua, TIAN Na
- 气候变化对我国黑胸大蠊分布的影响研究
- Effects of climate change on the distribution of Periplaneta fuliginosa in China
- 中国媒介生物学及控制杂志, 2023, 34(4): 542-547
- Chin J Vector Biol & Control, 2023, 34(4): 542-547
- 10.11853/j.issn.1003.8280.2023.04.018
-
文章历史
- 收稿日期: 2022-12-13



黑胸大蠊(Periplaneta fuliginosa)是重要的室内卫生害虫[1],能入侵人居住环境,较其他蜚蠊(也称蟑螂)更多分布在超市、医院、居民区等室内场所[2],会损坏室内物品、传播病原菌和寄生虫引起伤寒、霍乱等疾病、污染食物、造成过敏反应等[3],对人群日常生活和身体健康影响较大。
黑胸大蠊在我国分布广泛,目前主要分布于云南、贵州、四川、广西、广东、福建、浙江、江苏等省份[4],作为我国本土种类,被外来蜚蠊竞争后从优势种变为弱势种群,失去了部分生存环境,但蜚蠊监测显示部分地区黑胸大蠊比例在逐年升高[5],2018年后,徐州地区开始监测到黑胸大蠊,蠊种由单一到多样,密度也呈上升趋势[6]。同时,不同生境的不同蜚蠊种类携带的病原菌具有一定差异,如黄山市检测到仅黑胸大蠊携带鸡沙门菌(Salmonella ser. Gallinarum)[7]。虽然近年德国小蠊(Blattella germanica)逐渐发展为我国居民区蜚蠊优势种群,但黑胸大蠊在居民的生活场所中与人群的接触不可忽视,仍存在再次成为优势种和传播疾病的风险。
脏、乱、差的卫生环境和交通运输等人类活动会促进蜚蠊的孳生和传播,同时气候和地理因素变化与物种分布密切相关,黑胸大蠊的生存分布也受环境因素影响[8]。温度变化会使黑胸大蠊为寻找适宜的栖息地而发生频繁活动,在15~37 ℃黑胸大蠊随温度升高而活动增强[9]。在人类居住环境中,全球范围内的蜚蠊侵扰趋势在不断增加,影响人类相关疾病的患病率[10],故对我国未来黑胸大蠊分布区进行预测,据此进行针对性防控,有助于减少其侵扰和相关疾病传播。
最大熵模型(MaxEnt)可以利用物种分布数据与分布区域内环境变量数据,分析各变量与物种出现概率之间的关系,预测不同气候情景下的物种分布状态[11-12]。目前,运用MaxEnt模型预测环境因素对我国黑胸大蠊分布研究尚未见报道。本研究使用MaxEnt模型分析环境因素对我国黑胸大蠊分布的影响,并预测当前和未来气候情景下黑胸大蠊潜在适生区范围及适生区变化情况,为蜚蠊防治提供理论依据。
1 材料与方法 1.1 环境数据的获取与处理气候因子数据从全球气候数据网站(www.worldclim.org)下载,包括当前气候模式下19个气候因子,空间分辨率为5 arc min;下载共享社会经济路径(shared socio-economic pathway,SSP)2-4.5气候情景下,未来2041-2060年的19个气候因子(bio1~bio19)。自航天飞机雷达地形测绘任务网站(http://srtm.Csi.Cgiar.org/)获取海拔数据;利用ArcGIS 10.7软件表面分析功能获得坡度、坡向信息,一共获得温度、降水、海拔等环境因子22个。
1.2 黑胸大蠊分布点的获取与处理从已公开发表的文献、报告中检索“黑胸大蠊”或“Periplaneta fuliginosa”关键词,记录存在黑胸大蠊的位置信息,具体至县、区[13],通过百度地图的坐标拾取功能将分布地点转换为经纬度信息,同时从全球生物多样性信息交换平台(Global Biodiversity of Information Facility,GBIF)获取黑胸大蠊其他分布点信息。运用ArcGIS 10.7软件设置半径为5 km的缓冲区,与气候因子分辨率保持一致,去除重复、无效的分布点,使每个5 km×5 km栅格中仅保留1个分布点,以避免数据过于集中导致过拟合[14-15]。
在ArcGIS 10.7软件中,使用最邻近法重采样,使各环境数据空间分辨率一致,掩膜提取出中国部分。通过SPSS 25.0软件对环境因子进行相关性分析,避免因子间的高度共线性导致过度拟合[16]。将物种数据和环境数据导入maxent 3.4.4软件中分析,得到各个环境因子对模型的贡献率大小。当2个环境因子相关系数∣r∣≥0.9时[17],保留贡献率高的因子,最终筛选出13个环境因子纳入模型。
1.3 我国黑胸大蠊在当前气候模式下的潜在适生区预测随机选取70%的分布点作为训练集,剩余30%分布点作为测试集,交叉验证10次,构建MaxEnt模型,制作响应曲线。使用受试者工作特征(receiver operating characteristic,ROC)曲线下面积(area under curve,AUC)评价模型预测效果,AUC值> 0.8时认为模型预测结果可靠、精度良好[18]。
将MaxEnt模型的预测结果导入ArcGIS 10.7软件,获得我国黑胸大蠊潜在适生区分布图,采用重分类中“自然间断点分级法”,按照分布概率划分4个适生区等级:< 0.12为非适生区,0.12~0.31(不含0.31)为低适生区,0.31~0.50为中适生区,> 0.50为高适生区。
1.4 我国黑胸大蠊在未来气候模式下的适生区变化预估将筛选出的13个环境因子的未来环境数据与当前环境数据一一对应输入MaxEnt模型中,预估未来黑胸大蠊的潜在分布区。将当前与未来的预测结果导入ArcGIS 10.7软件中,借助Spatial Analyst工具的组合或功能获得当前和未来适生区分布变化情况。
2 结果 2.1 黑胸大蠊分布点经检索共获取有关黑胸大蠊中文文献1 442篇、外文文献222篇。从文献中共收集黑胸大蠊物种分布点397个,从GBIF中收集16个物种分布点,缓冲区筛选后最终剩余259个分布点。
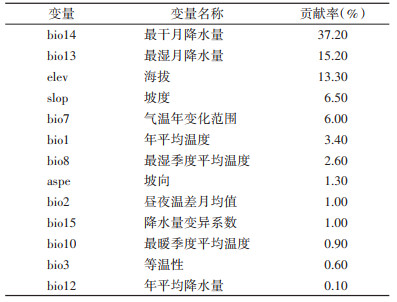
2.2 环境因子对黑胸大蠊分布的影响13个环境因子用于构建MaxEnt模型,最干月降水量(bio14)对黑胸大蠊分布的影响最大,贡献率达37.20%,其次为最湿月降水量(15.20%)、海拔(13.30%)。见表 1。
|
贡献率排在前3位的3种环境因子响应曲线见图 1。结果表明,降水量对黑胸大蠊分布影响较大,随最干月降水量的升高,黑胸大蠊的出现概率先上升后缓慢下降。最干月降水量达15.09 mm时出现概率超过0.50;最干月降水量处于33.46 mm时,出现概率达到最高。

|
| 图 1 黑胸大蠊出现概率对重要环境因子的响应曲线 Figure 1 Response curves of Periplaneta fuliginosa occurrence probability to major environmental factors |
| |
随着最湿月降水量(bio13)的升高,黑胸大蠊的出现概率先上升后下降。最湿月降水量在168.38~292.41 mm时,出现概率 > 0.50;在177.62 mm时,出现概率最高,之后随最湿月降水量升高黑胸大蠊出现概率降低。
随着海拔(elev)的升高,黑胸大蠊的出现概率下降,海拔在105.16 m时,出现概率 < 0.50。
2.3 当前气候条件下我国黑胸大蠊的潜在分布区模型验证结果表明,ROC曲线下面积为0.9,提示模型预测精度较好。预测结果显示,当前气候条件下黑胸大蠊的高、中、低适生区面积分别为61.31、106.57和105.39万km2(表 2)。其中,高适生区主要分布在安徽、江苏、江西等地区。见图 2。

|
| 图 2 当前气候情景下黑胸大蠊的潜在适生区及物种采样点 Figure 2 Potential suitable areas and sampling sites of Periplaneta fuliginosa under current climate conditions |
| |
SSP2-4.5气候情景下,未来2041-2060年全国黑胸大蠊总适生区面积将减少4.29万km2,新增适生区面积1.47万km2,损失适生区位于西藏、云南、四川等地区,新增适生区位于陕西、山西、吉林、台湾等地区(图 3)。其中,高适生区面积将减少2.96万km2;中适生区面积减少2.99万km2;低适生区面积增加1.66万km2;非适生区面积增加4.29万km2。见表 2。

|
| 图 3 当前和2041-2060年共享社会经济路径2-4.5气候情景下黑胸大蠊适生区分布变化 Figure 3 Changes of suitable areas of Periplaneta fuliginosa under current conditions and the shared socioeconomic pathway SSP2-4.5 scenario in 2041-2060 |
| |

|
本研究应用MaxEnt模型,分析了环境因子对我国黑胸大蠊分布区的影响,并对当前及未来气候情景下黑胸大蠊的分布区进行了预测。结果表明,黑胸大蠊主要分布于温暖潮湿的南方地区,最干月降水量、最湿月降水量、海拔是影响黑胸大蠊分布的主要环境因子,其中最干月降水量贡献率最大,达37.20%,坡度、气温年变化范围、年平均温度等环境因子也会影响其分布。戈斌等[19]对上海市奉贤区蜚蠊监测发现蜚蠊密度与气温、湿度、降水量之间呈正相关,在7、8月梅雨季节时,蜚蠊密度达到高峰。黑胸大蠊的室内繁殖研究中[20],其卵孵受温度、湿度影响较为明显,3-8月时卵孵率较高,而低温、干燥的秋冬季节不适宜黑胸大蠊发育繁殖。福建省宁德市山区县海拔300~900 m,黑胸大蠊密度随海拔增高而增大,与本研究结果有差异,但沿海县海拔50~100 m,密度比山区县更大[21],考虑为湿度对黑胸大蠊分布比海拔影响更大所致。
物种出现概率≥0.5所对应的环境因子范围是物种的高适生区域[22]。据研究可知,蜚蠊喜欢选择温暖、潮湿、食物丰富和多缝隙的隐蔽场所栖居,这是它们孳生繁殖所需要的4个基本条件。黑胸大蠊为亚热带蜚蠊种类,在我国分布于云南、贵州、广东、福建、江苏等省份,本研究预测的黑胸大蠊潜在适生区分布与其实际分布相符。黑胸大蠊高适生区分布的上海、江浙等地区,大多属于亚热带季风气候,温暖湿润,最干月降水量在10~50 mm,最湿月降水量在200 mm左右,适宜黑胸大蠊生长,终年有蜚蠊活动,而最干月降水量 < 10 mm的新疆、西藏等西北部城市不适宜其生存,分布较少。同时,预测显示黑胸大蠊适生区主要为平原、丘陵、盆地地形,海拔基本在200 m左右,符合环境因子对其影响研究得出的黑胸大蠊适宜生存的海拔条件,文献报道浙江省在低海拔地区蜚蠊种类、数量更多[23],也验证了这一结论。
在SSP2-4.5气候情景下,未来2041-2060年全国黑胸大蠊总体上适生区呈轻微减少状态,研究结果显示,黑胸大蠊出现概率先随降水量增加而增加,当最干月降水量超过33.46 mm或最湿月降水量超过177.62 mm后,出现概率有所下降。据研究预测[24-25],未来2041-2060年,我国降水增加,雨带季节延长,华南地区夏季降雨增量达到1 mm/d,大量的降水可能是黑胸大蠊适生区总面积减少的原因。适生区变化的敏感范围位于西藏、云南、四川、陕西、山西、辽宁、吉林、黑龙江等省份,国家气候中心气候系统模式数据显示这些地区年降水量在400~800 mm,该区域黑胸大蠊易随降水量等环境因子变化而导致适生区等级以及适生面积发生变化。台湾省的适生区变化较为特殊,适生区分布主要集中在台湾岛沿海区域,未来山脉西部的一些适生区将会由非适生区转为适生区,这可能受台湾省内山脉及坡度影响。
预防黑胸大蠊造成过敏性鼻炎或哮喘等致敏性疾病,传播肠道致病菌、病毒,应对室内黑胸大蠊进行监测和防制,可根据最干月降水量、最湿月降水量的变化判断黑胸大蠊侵害季节消长规律,选择适当的防制时机,采取有效防制措施,海拔105.16 m以下的黑胸大蠊高出现概率地区以及预测的未来气候情景下黑胸大蠊分布的高适生区及敏感区域,将为决策防制地区和范围的划分提供科学依据。
本研究选用的是气候、地形有关的环境变量,缺少人类社会活动对蜚蠊分布可能造成的影响,但人类活动因素较为复杂,本研究限于相关专业知识和数据的可获得性,无法全面评估人群活动变化情况。未来可以尝试纳入更多影响因素以更准确地研究黑胸大蠊的分布、预测。
利益冲突 无
| [1] |
贺盼, 马强. 我国蜚蠊的分布及防制概况[J]. 医学动物防制, 2018, 34(9): 868-872. He P, Ma Q. Distribution and control of cockroaches in China[J]. J Med Pest Control, 2018, 34(9): 868-872. |
| [2] |
侯银续, 林浩飞, 王玮, 等. 安徽省黄山市2017-2018年室内蜚蠊监测结果分析[J]. 中国媒介生物学及控制杂志, 2020, 31(6): 718-721. Hou YX, Lin HF, Wang W, et al. An analysis of surveillance results of indoor cockroaches in Huangshan, Anhui province, China, 2017-2018[J]. Chin J Vector Biol Control, 2020, 31(6): 718-721. DOI:10.11853/j.issn.1003.8280.2020.00.020 |
| [3] |
侯银续, 水岩, 吴磊, 等. 蜚蠊的危害及防制技术研究进展[J]. 安徽预防医学杂志, 2021, 27(6): 487-491, 496. Hou YX, Shui Y, Wu L, et al. Research progress on cockroach damage and control technology[J]. Anhui J Prev Med, 2021, 27(6): 487-491, 496. DOI:10.19837/j.cnki.ahyf.2021.06.017 |
| [4] |
孙耘芹, 李梅, 何凤琴, 等. 五种蜚蠊的生物学特性和综合治理[J]. 昆虫知识, 2004, 41(3): 216-222. Sun YQ, Li M, He FQ, et al. Bionomics and integrated management of five species of cockroaches[J]. Entomol Knowl, 2004, 41(3): 216-222. DOI:10.3969/j.issn.0452-8255.2004.03.006 |
| [5] |
王韶华, 武峥嵘, 董腾飞, 等. 上海市嘉定区2017-2019年蟑螂侵害现状及居家灭蟑效果研究[J]. 上海预防医学, 2020, 32(12): 1001-1005. Wang SH, Wu ZR, Dong TF, et al. Surveillance of cockroach population and evaluation of control effects in households from 2017 to 2019 in Jiading district of Shanghai[J]. Shanghai J Prev Med, 2020, 32(12): 1001-1005. DOI:10.19428/j.cnki.sjpm.2020.20324 |
| [6] |
刘雪, 葛小伍, 梁丽, 等. 2017―2019年徐州市主城区主要病媒生物监测分析[J]. 中华卫生杀虫药械, 2021, 27(2): 124-128. Liu X, Ge XW, Liang L, et al. Monitoring and analysis of the main vectors in main districts of Xuzhou from 2017 to 2019[J]. Chin J Hyg Insect Equip, 2021, 27(2): 124-128. DOI:10.19821/j.1671-2781.2021.02.009 |
| [7] |
侯银续, 袁华玲, 陈李, 等. 黄山市蟑螂携带肠道菌种多样性初探[J]. 中华卫生杀虫药械, 2021, 27(2): 154-159. Hou YX, Yuan HL, Chen L, et al. A preliminary study on the diversity of intestinal bacteria carried by cockroaches in Huangshan city[J]. Chin J Hyg Insect Equip, 2021, 27(2): 154-159. DOI:10.19821/j.1671-2781.2021.02.017 |
| [8] |
赵泽芳, 卫海燕, 郭彦龙, 等. 人参潜在地理分布以及气候变化对其影响预测[J]. 应用生态学报, 2016, 27(11): 3607-3615. Zhao ZF, Wei HY, Guo YL, et al. Potential distribution of Panax ginseng and its predicted responses to climate change[J]. Chin J Appl Ecol, 2016, 27(11): 3607-3615. DOI:10.13287/j.1001-9332.201611.040 |
| [9] |
姜志宽, 吴光华. 蟑螂防治(二): 蟑螂的生态习性与常见种类[J]. 中华卫生杀虫药械, 2009, 15(2): 169-172. Jiang ZK, Wu GH. Cockroach control. Ⅱ: Ecological habits and common species of cockroaches[J]. Chin J Hyg Insect Equip, 2009, 15(2): 169-172. DOI:10.19821/j.1671-2781.2009.02.032 |
| [10] |
Nasirian H. Infestation of cockroaches (Insecta: Blattaria) in the human dwelling environments: A systematic review and meta-analysis[J]. Acta Trop, 2017, 167: 86-98. DOI:10.1016/j.actatropica.2016.12.019 |
| [11] |
张路. MaxEnt最大熵模型在预测物种潜在分布范围方面的应用[J]. 生物学通报, 2015, 50(11): 9-12. Zhang L. The application of MaxEnt model in predicting the potential range of species[J]. Biol Bull, 2015, 50(11): 9-12. |
| [12] |
Besprozvannykh VV, Ngo HD, Ha NV, et al. Descriptions of digenean parasites from three snail species, Bithynia fuchsiana (Morelet), Parafossarulus striatulus Benson and Melanoides tuberculata Müller, in North Vietnam[J]. Helminthologia, 2013, 50(3): 190-204. DOI:10.2478/s11687-013-0131-5 |
| [13] |
Ren ZP, Wang DQ, Ma AM, et al. Predicting malaria vector distribution under climate change scenarios in China: Challenges for malaria elimination[J]. Sci Rep, 2016, 6(1): 20604. DOI:10.1038/srep20604 |
| [14] |
王茹琳, 李庆, 封传红, 等. 基于MaxEnt的西藏飞蝗在中国的适生区预测[J]. 生态学报, 2017, 37(24): 8556-8566. Wang RL, Li Q, Feng CH, et al. Predicting potential ecological distribution of Locusta migratoria tibetensis in China using MaxEnt ecological niche modeling[J]. Acta Ecol Sin, 2017, 37(24): 8556-8566. DOI:10.5846/stxb201611152326 |
| [15] |
周玉婷, 葛雪贞, 邹娅, 等. 基于MaxEnt模型的长林小蠹的全球及中国适生区预测[J]. 北京林业大学学报, 2022, 44(11): 90-99. Zhou YT, Ge XZ, Zou Y, et al. Prediction of the potential geographical distribution of Hylurgus ligniperda at the global scale and in China using the MaxEnt model[J]. J Beijing For Univ, 2022, 44(11): 90-99. DOI:10.12171/j.1000-1522.20210345 |
| [16] |
曾权, 朱雪珍, 周利娟. 基于优化MaxEnt模型的南方三棘果在中国的潜在适生区预测[J]. 华南农业大学学报, 2023, 44(2): 254-262. Zeng Q, Zhu XZ, Zhou LJ. Prediction of potential suitable region for Emex australis in China based on the optimized MaxEnt mode[J]. J South China Agric Univ, 2023, 44(2): 254-262. |
| [17] |
Wouyou HG, Lokonon BE, Idohou R, et al. Predicting the potential impacts of climate change on the endangered Caesalpinia bonduc (L.) Roxb in Benin (West Africa)[J]. Heliyon, 2022, 8(3): e09022. DOI:10.1016/j.heliyon.2022.e09022 |
| [18] |
Liu D, Lei XD, Gao WQ, et al. Mapping the potential distribution suitability of 16 tree species under climate change in northeastern China using MaxEnt modelling[J]. J For Res, 2022, 33(6): 1739-1750. DOI:10.1007/s11676-022-01459-4 |
| [19] |
戈斌, 黄涛, 张海兵, 等. 2011-2018年上海市奉贤区蟑螂监测分析[J]. 中华卫生杀虫药械, 2021, 27(5): 425-428. Ge B, Huang T, Zhang HB, et al. Investigation on cockroaches in Fengxian district of Shanghai from 2011 to 2018[J]. Chin J Hyg Insect Equip, 2021, 27(5): 425-428. DOI:10.19821/j.1671-2781.2021.05.010 |
| [20] |
徐绍锐, 汪世平, 吴仕筠, 等. 黑胸大蠊室内繁殖、发育的生物学特性研究[J]. 中国人兽共患病学报, 2006, 22(10): 913-917. Xu SR, Wang SP, Wu SJ, et al. Studies on biological characteristics of domestic reproduction and development of Periplaneta fuliginosa[J]. Chin J Zoonoses, 2006, 22(10): 913-917. DOI:10.3969/j.issn.1002-2694.2006.10.005 |
| [21] |
陈文锦, 袁高林. 宁德市蜚蠊种群密度与季节消长[J]. 海峡预防医学杂志, 2006, 12(6): 28-29. Chen WJ, Yuan GL. Population density and seasonal fluctuation of cockroaches in Ningde city[J]. Strait J Prev Med, 2006, 12(6): 28-29. DOI:10.3969/j.issn.1007-2705.2006.06.014 |
| [22] |
李芮芝, 胡希军, 文亚峰, 等. 基于MaxEnt模型的丹霞梧桐生境预测模拟分析[J]. 中国野生植物资源, 2022, 41(6): 1-7, 30. Li RZ, Hu XJ, Wen YF, et al. Habitat prediction simulation analysis of Firmiana danxiaensis based on MaxEnt[J]. Chin Wild Plant Resour, 2022, 41(6): 1-7, 30. DOI:10.3969/j.issn.1006-9690.2022.06.001 |
| [23] |
沈培谊, 俞松青, 蔡昌曜, 等. 浙江省蜚蠊种群、地理分布特点及防制技术研究[J]. 中国媒介生物学及控制杂志, 2001, 12(4): 275-281. Shen PY, Yu SQ, Cai CY, et al. The study on the features of populations and distribution and control technique of the cockroach in Zhejiang province[J]. Chin J Vector Biol Control, 2001, 12(4): 275-281. DOI:10.3969/j.issn.1003-4692.2001.04.010 |
| [24] |
姜荣升. 基于CWRF研究我国降水变化及其与边界层高度的关系[D]. 南京: 南京信息工程大学, 2021. DOI: 10.27248/d.cnki.gnjqc.2021.000035. Jiang RS. Study the interaction between precipitation change and planetary boundary layer height based on CWRF[D]. Nanjing: Nanjing University of Information Science and Technology, 2021. DOI: 10.27248/d.cnki.gnjqc.2021.000035.(in Chinese) |
| [25] |
刘春蓁, 巢清尘, 王守荣, 等. 水文气象学领域的水文循环研究进展[J]. 气候变化研究进展, 2023, 19(1): 1-10. Liu CZ, Chao QC, Wang SR, et al. The research progress of hydrologic cycle in hydrometeorology[J]. Climate Change Res, 2023, 19(1): 1-10. DOI:10.12006/j.issn.1673-1719.2022.174 |