 2023, Vol. 34
2023, Vol. 34扩展功能
文章信息
- 刘雨秋, 鲁亮, 刘蓬勃, 赵宁, 李贵昌, 栗冬梅, 宋秀平, 王君, 刘起勇
- LIU Yu-qiu, LU Liang, LIU Peng-bo, ZHAO Ning, LI Gui-chang, LI Dong-mei, SONG Xiu-ping, WANG Jun, LIU Qi-yong
- 内蒙古自治区3个地区布氏田鼠的种群遗传多样性及种群遗传结构研究
- Genetic diversity and genetic structure of Lasiopodomys brandtii populations in three regions of Inner Mongolia, China
- 中国媒介生物学及控制杂志, 2023, 34(3): 291-297
- Chin J Vector Biol & Control, 2023, 34(3): 291-297
- 10.11853/j.issn.1003.8280.2023.03.002
-
文章历史
- 收稿日期: 2023-01-05




布氏田鼠(Lasiopodomys brandtii)隶属于啮齿目仓鼠科田鼠亚科,主要分布在东北亚草原地带,包括我国内蒙古自治区(内蒙古)中部和东部部分地区,河北省北部的局部地区,以及蒙古共和国南部和俄罗斯。在我国布氏田鼠分布于内蒙古中部锡林郭勒草原、东部呼伦贝尔草原及河北省北部的张北草原,其中锡林郭勒草原、呼伦贝尔草原是其主要分布区。布氏田鼠在内蒙古草原的分布是不连续的,呈间断式、片段式分布[1]。布氏田鼠是典型的植食性鼠类,通过挖掘家族聚居的洞系,啃食草根、幼苗,破坏草场,导致草原沙漠化,使其向着不利于生态系统的方向发展。大面积高密度的布氏田鼠啃食牧草,严重破坏牧场,导致牛、羊群搬迁,影响畜牧业的发展,给人类经济带来巨大损失[2-4]。近年来,放牧、开垦草地、采挖药材等人类活动导致的草原生态系统退化[5],以及气候、栖息地、生境的变化等都会引起布氏田鼠的分布和迁移[5]。同时,布氏田鼠是锡林郭勒高原布氏田鼠鼠疫自然疫源地的重要宿主动物。该疫源地分布于内蒙古锡林郭勒盟阿巴嘎熔岩台地和乌珠穆沁盆地西部一带,是内蒙古4类鼠疫疫源地之一[6]。1970-1975年在该地区出现了布氏田鼠间鼠疫的暴发流行,其中布氏田鼠占染疫动物的84%[7]。内蒙古草原在19世纪90年代至2010年代发生过布氏田鼠的暴发,但具体的种群动态调节机制目前尚不清楚[8-9]。
布氏田鼠的数量变化有一定的规律,主要分为上升期、高峰期和下降期,一般上升期1~2年,下降期1~2年,低谷期一般10年以上[10]。布氏田鼠主要在上升期和高峰期对草场造成危害[11];其种群密度受内在和外在因素的驱动,时常会发生较大的波动[11]。在低谷期,分布区南部的布氏田鼠密度非常低,很难监测到;但在上升期则很快形成局部暴发。这些形成暴发的布氏田鼠种群是在本地长期存在,还是由外地迁徙而来尚无明确的解释。因此,分析布氏田鼠的种群遗传多样性和种群遗传结构对于了解布氏田鼠种群的迁徙和波动具有重要的实践意义。
DNA分子标记法是指可以遗传且能够检测的DNA序列,是个体的遗传物质也是核苷酸序列的基础,能够从分子水平分析个体遗传变异和遗传分化[12]。微卫星标记又称短串联重复序列、简单重复序列,一般是由2~6个碱基组成的核苷酸序列,为共显性标记,能够稳定遗传,而且容易获得,结果重复性高,广泛用于动物遗传多样性研究[13-15]。姜姗等[16]用微卫星标记的方法分析了我国东北地区灰飞虱(Laodelphax striatellus)的种群遗传结构,得出在不同地区灰飞虱存在一定程度的遗传分化,且基因交流较低,存在一定的种群遗传结构差异的结论。康芬芬等[17]用此方法分析橘小实蝇(Bactrocera dorsalis)4个地理种群的遗传多态性,结果显示橘小实蝇4个地理种群存在一定的遗传差异,地理隔离是造成差异的主要原因。本研究通过微卫星标记的方法,分析比较内蒙古3个样地和不同月份布氏田鼠的种群遗传多样性和种群遗传结构,为进一步分析布氏田鼠不同地理种群在种群数量动态变化过程中的种群遗传特征提供科学依据。
1 材料与方法 1.1 样本来源2021年5、7、9月,使用鼠夹法和鼠笼法采集我国内蒙古新巴尔虎右旗(HL)、东乌珠穆沁旗(XM)和锡林浩特市(QL)3个样地的布氏田鼠,通过形态学鉴定物种,解剖布氏田鼠取肝脏组织,保存于-80 ℃冰箱。
1.2 方法 1.2.1 微卫星位点获取微卫星分析所用引物主要参考已发表文献[18-21],共选择16对扩增结果稳定、扩增条带清晰的引物对布氏田鼠进行种群遗传分析。16对引物正向5′端分别用马来酰亚胺(ROX,红色)、羧基荧光素(FAM,蓝色)、六氯荧光素(HEX,绿色)、羧基四甲基罗丹明(TAMRA,黄色)4种不同的荧光基团修饰。
1.2.2 微卫星位点的PCR扩增根据磁珠法DNA提取试剂盒说明书,取布氏田鼠肝脏,提取每只鼠的DNA,按照16对荧光标记的正、反向引物进行PCR扩增。PCR扩增的反应体系:1-5TM2X High-Fidelity Master Mix 12.5 μl,Water ddH2O 9.5 μl,Template DNA 1.0 mmol/μl,Primer F 1.0 μl,Primer R 1.0 μl。PCR扩增的反应条件:95 ℃预变性5 min;95 ℃变性30 s,根据不同的引物设置退火温度,72 ℃延伸30 s,共30个循环;95 ℃延伸30 s,4 ℃保存。阳性扩增产物进行毛细管电泳分析,结果再用GeneMarker软件获得微卫星片段长度数据。
1.2.3 分析数据利用MS-Tool进行微卫星数据的录入和数据异常的纠错。利用GenAIEx 6.5软件计算样本量(n)、平均等位基因数(Na)、有效等位基因数(Ne)、观察杂合度(Ho)、期望杂合度(He)、无偏期望杂合度(uHe)和香农信息指数(Shannon’s information index,I),利用PIC-CALC软件计算多态信息量(PIC)。利用structure 2.3.4软件计算聚类数K值[22],分析布氏田鼠种群遗传结构,基于遗传距离(GD)和地理距离(GGD)用GenAIEx 6.5软件做主成分分析。利用Arlequin(version 30)软件进行种群的分子方差分析(AMOVA)。
2 结果 2.1 布氏田鼠的种群遗传多样性用16个微卫星位点对内蒙古3个样地3个月的9个样本群总计275只布氏田鼠进行微卫星位点分析,共检测到472个等位基因。所有位点均具有多态性,各微卫星位点平均等位基因数波动区间为6.222~17.222,16个位点等位基因平均值为10.264。16个微卫星位点的多态信息量波动区间为0.579 6~0.938 6,平均值为0.821 3。16个微卫星位点的香农信息指数波动范围为1.192~2.504,平均值为1.812。16个微卫星位点的观察杂合度波动区间为0.478~0.951,期望杂合度的波动区间为0.575~0.891,这5个指标均是反映种群遗传多样性的指数,数值越大,代表遗传多样性越高,遗传信息越丰富(表 1)。基于不同样地和不同月份的布氏田鼠样本群的遗传多样性结果显示:9个样本群的每个位点平均等位基因数波动范围为8.813~12.188,平均值为10.264。9个样本群的香农信息指数波动范围为1.691~2.046,平均值为1.812。各样本群的观察杂合度波动范围为0.585~0.823,期望杂合度的波动范围为0.734~0.816。见表 2、图 1。
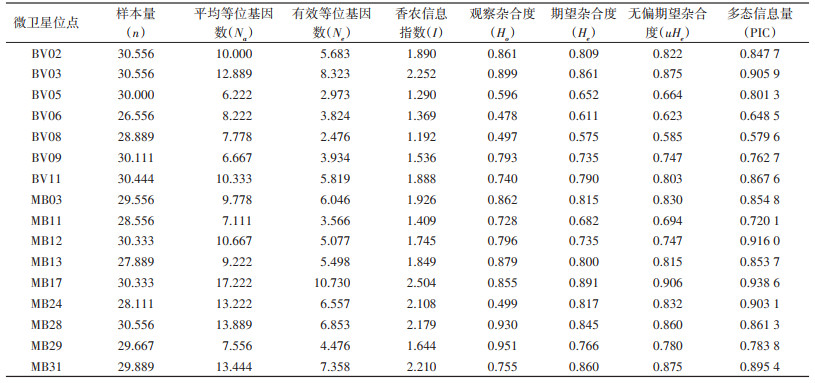
|

|

|
| 注:HLM新巴尔虎右旗(2021年5月);XMM东乌珠穆沁旗(2021年5月);QLM锡林浩特市(2021年5月);HLJ新巴尔虎右旗(2021年7月);XMJ东乌珠穆沁旗(2021年7月);QLJ锡林浩特市(2021年7月);HLS新巴尔虎右旗(2021年9月);XMS东乌珠穆沁旗(2021年9月);QLS锡林浩特市(2021年9月);图注括号内为标本采集时间。 图 1 基于样地的布氏田鼠样本群遗传多样性 Figure 1 Genetic diversity of Lasiopodomys brandtii populations based on sample plots |
| |
利用structure 2.3.4软件分析3个样地3个月布氏田鼠的样本群遗传结构,根据DeltaK法确定最优类群数K值等于4,提示9个样本群共275只布氏田鼠可以分为4个不同的类群,4个类群分别为2021年5月新巴尔虎右旗、东乌珠穆沁旗、锡林浩特市(类群Ⅰ,橙色);2021年7月新巴尔虎右旗、东乌珠穆沁旗、锡林浩特市(类群Ⅱ,粉色);2021年9月新巴尔虎右旗、锡林浩特市(类群Ⅲ,蓝色);2021年9月东乌珠穆沁旗(类群Ⅳ,黄色)。4个类群整体上可以分开,界限较清晰,部分混入的个体可以解释为渗入。4个类群之间的渗入是不对称的,类群Ⅲ流向类群Ⅱ > 类群Ⅳ,类群Ⅱ流向类群Ⅲ > 类群Ⅰ,类群Ⅲ流向类群Ⅰ > 类群Ⅳ。聚类分析结果与GenAIEx 6.5软件主成分分析结果一致。见图 2、3。

|
| 注:HLM新巴尔虎右旗(2021年5月);XMM东乌珠穆沁旗(2021年5月);QLM锡林浩特市(2021年5月);HLJ新巴尔虎右旗(2021年7月);XMJ东乌珠穆沁旗(2021年7月);QLJ锡林浩特市(2021年7月);HLS新巴尔虎右旗(2021年9月);XMS东乌珠穆沁旗(2021年9月);QLS锡林浩特市(2021年9月);以上均为布氏田鼠的样本种群,括号内为标本采集时间。 图 2 布氏田鼠样本群聚类结果 Figure 2 Clustering results of Lasiopodomys brandtii populations |
| |

|
| 注:HLM新巴尔虎右旗(2021年5月);XMM东乌珠穆沁旗(2021年5月);QLM锡林浩特市(2021年5月);HLJ新巴尔虎右旗(2021年7月);XMJ东乌珠穆沁旗(2021年7月);QLJ锡林浩特市(2021年7月);HLS新巴尔虎右旗(2021年9月);XMS东乌珠穆沁旗(2021年9月);QLS锡林浩特市(2021年9月);以上均为布氏田鼠的样本种群,括号内为标本采集时间。 图 3 布氏田鼠的主成分分析图 Figure 3 Principal component analysis map of Lasiopodomys brandtii |
| |
根据布氏田鼠种群遗传结构,将9个布氏田鼠样本群按照4个类群进行分子方差分析,结果见表 3。4个类群间的遗传变异占总变异的5.04%,类群种群间、种群间个体分别占总变异的3.89%和2.56%。个体间的变异占总变异的88.51%。
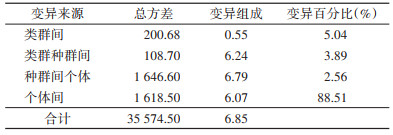
|
根据GenAIEx 6.5软件中的mantel test检验遗传距离和地理距离的相关性,通过GenAIEx 6.5软件中的creat输入每个样地的经纬度计算地理距离,结果显示遗传距离与地理距离无显著相关性。见图 4。

|
| 图 4 遗传距离和地理距离的相关关系 Figure 4 Correlation between genetic distance and geographical distance |
| |
遗传分化系数(FST)是评价种群遗传分化的主要指标,FST在0~0.05,种群之间遗传分化程度较低,FST在0.05~0.15,种群间遗传分化中等,FST在0.15~0.25,种群遗传分化水平较高,FST > 0.25时,种群间遗传分化程度极高。本次研究基于16对荧光标记引物,分析中国内蒙古地区9个布氏田鼠样本群的遗传分化表明9个样本群之间的FST区间是0.017~0.088,其中XMM(2021年5月东乌珠穆沁旗)和QLM(2021年5月锡林浩特市)2个样本群的遗传分化系数最低(FST=0.017),HLM(2021年5月新巴尔虎右旗)和XMS(2021年9月东乌珠穆沁旗)2个样本群的FST最高(FST=0.088)。成对的FST中,有44.44%的FST < 0.05,有55.56%的FST在0.05~0.15。
此次研究的9个样本群的基因流(Nm)区间为2.604~14.109,其中QLM和XMM样本群之间的Nm最大(Nm=14.109),XMS和HLM样本群之间的Nm最小(Nm=2.604)。所有样本群间的Nm均 > 1。见表 4。

|
通过查阅相关文献,本文利用16个微卫星标记于2021年5、7、9月在内蒙古新巴尔虎右旗、东乌珠穆沁旗和锡林浩特市3个样地对布氏田鼠进行样本群遗传多样性和样本群遗传结构分析。每个微卫星位点最少有6个等位基因,最多达到17个。PIC是反映样本群遗传多样性的较好指标,当PIC > 0.5时,认为该位点具有高度多态性[23],因此,本研究所选16个微卫星位点均具有高度多态性(均 > 0.5)。香农信息指数和PIC的结果一致,即香农信息指数高的位点,PIC值也高。位点遗传多样性指标越高,能提供的遗传信息越多,这有利于做种群遗传多样性分析。当期望杂合度 > 0.5时,认为该种群具有较高的遗传多样性[24]。结合本次布氏田鼠的观察杂合度、期望杂合度、PIC、香农信息指数等种群遗传多样性指标,16个微卫星标记能够反映内蒙古地区布氏田鼠的种群遗传多样性,可以用于布氏田鼠的种群遗传研究。
基于structure聚类分析结果,9个布氏田鼠的样本群聚为4类,主要呈现月份间的差异。布氏田鼠的样本群遗传结构呈现明显的季节性,这种季节性结构称为每年的繁殖季节和非繁殖季节的种群遗传结构[25]。计算4个类群的布氏田鼠的种群遗传多样性,结果显示,等位基因数、有效等位基因数、观察杂合度、期望杂合度和香农信息指数等反映遗传多样性指标均为5月> 7月> 9月,即布氏田鼠的种群遗传多样性春季 > 夏季 > 秋季,该结果符合“增殖-扩散-平衡”假说[26],该假说认为,在一定的自然条件下,由于繁殖季节布氏田鼠一直在产生新的个体,个体间的亲缘关系较近,所以种群的遗传多样性较低,在非繁殖季节,没有新生个体的不断补充,只有扩散或迁移,布氏田鼠个体间的亲缘关系较远,种群的遗传多样性较高。关于种群遗传距离与地理距离的mantel test相关性检验,该结果与Wang等[27]用7个微卫星位点对内蒙古呼伦贝尔分布区、锡林郭勒北部分布区和浑善达克沙地南部分布区等4个样地共158只布氏田鼠的mantel test结果一致(r=0.049 9,P=0.669),即各种群间遗传距离和地理距离无显著相关性。
本研究的9个内蒙古布氏田鼠样本群FST值均 < 0.15,说明2021年内蒙古3个地区的布氏田鼠种群处于中度及低度分化阶段。Nm是指一个群体向另一个群体产生的基因交换的总称[28],即部分个体从一个群体迁移到另一个群体,从而把新的基因带到另一个群体中,产生的基因流动,Nm值越大,说明群体间的基因交流越频繁,群体间个体的差异越小,群体间的相似度越大[29]。此次9个样本群中XMM(2021年5月东乌珠穆沁旗)和QLM(2021年5月锡林浩特市)2个样本群间的Nm值最大(Nm=14.109),说明这2个样本群之间的基因交流频繁。
利益冲突 无
| [1] |
蒋永恩, NanjE, BatsaikhanN, 等. 布氏田鼠栖息地特征的研究[J]. 草地学报, 2012, 20(1): 179-182. Jiang YE, Nanj E, Batsaikhan N, et al. Study on the habitat characteristics of Brandt's vole[J]. Acta Agrestia Sin, 2012, 20(1): 179-182. DOI:10.3969/j.issn.1007-0435.2012.01.028 |
| [2] |
Bai DF, Wan XR, Li GL, et al. Factors influencing range contraction of a rodent herbivore in a steppe grassland over the past decades[J]. Ecol Evol, 2022, 12(2): e8546. DOI:10.1002/ece3.8546 |
| [3] |
Zhang ZB, Pech R, Davis S, et al. Extrinsic and intrinsic factors determine the eruptive dynamics of Brandt's voles Microtus brandti in Inner Mongolia, China[J]. Oikos, 2003, 100(2): 299-310. DOI:10.2307/3548187 |
| [4] |
王涛涛, 白玛次仁, 海淑珍, 等. 布氏田鼠行为谱的构建[J]. 草地学报, 2015, 23(3): 646-652. Wang TT, Bai MCT, Hai SZ, et al. Construction for the ethogram of Brandt's voles (Lasiopodomys brandtii)[J]. Acta Agrestia Sin, 2015, 23(3): 646-652. DOI:10.11733/j.issn.1007-0435.2015.03.030 |
| [5] |
李素英, 李晓兵, 王丹丹. 基于马尔柯夫模型的内蒙古锡林浩特典型草原退化格局预测[J]. 生态学杂志, 2007, 26(1): 78-82. Li SY, Li XB, Wang DD. Prediction of grassland degradation in Xilinhaote of Inner Mongolia based on Markov process model[J]. Chin J Ecol, 2007, 26(1): 78-82. |
| [6] |
杨普伊, 李建云, 张喆. 内蒙古自治区不同类型鼠疫自然疫源地土壤金属元素含量调查分析[J]. 中国媒介生物学及控制杂志, 2022, 33(6): 900-905. Yang PY, Li JY, Zhang Z. Analysis of the content of metal elements in soil of different types of natural plague foci in Inner Mongolia Autonomous Region, China[J]. Chin J Vector Biol Control, 2022, 33(6): 900-905. DOI:10.11853/j.issn.1003.8280.2022.06.025 |
| [7] |
樊振亚. 布氏田鼠鼠疫对人危害性的初步分析[J]. 中华流行病学杂志, 1986, 7(6): 376-377. Fan ZY. Preliminary analysis of the harm of Lasiopdomys brandtii plague to human[J]. Chin J Epidemiol, 1986, 7(6): 376-377. DOI:10.3760/cma.j.issn.0254-6450.1986.06.136 |
| [8] |
张亚楠, 赵钢, 张大禹, 等. 2001-2013年内蒙古布氏田鼠鼠疫自然疫源地鼠疫监测结果分析[J]. 中华地方病学杂志, 2015, 34(10): 765-768. Zhang YN, Zhao G, Zhang DY, et al. Analysis of monitoring results on plague of rats in Lasiopodomys brandti plague natural foci in Inner Mongolia from 2001 to 2013[J]. Chin J Endemiol, 2015, 34(10): 765-768. DOI:10.3760/cma.j.issn.2095-4255.2015.10.016 |
| [9] |
Yin BF, Li GL, Wan XR, et al. Large manipulative experiments reveal complex effects of food supplementation on population dynamics of Brandt's voles[J]. Sci China Life Sci, 2017, 60(8): 911-920. DOI:10.1007/s11427-017-9114-9 |
| [10] |
董维惠, 侯希贤, 杨玉平. 掌握草原害鼠数量变动规律开展综合防治[J]. 中国草地, 2003, 25(6): 41-44, 62. Dong WH, Hou XX, Yang YP. The grasping on the law of population dynamics of rodents and carrying out the integrated control measures[J]. Grassland China, 2003, 25(6): 41-44, 62. DOI:10.3969/j.issn.1673-5021.2003.06.010 |
| [11] |
董维惠, 侯希贤, 杨玉平, 等. 草原和农田几种主要鼠种数量动态研究及预测[J]. 中国草地学报, 2008, 30(5): 90-95. Dong WH, Hou XX, Yang YP, et al. Studies on the population dynamics and prediction of several predominant rodents on grassland and farmland[J]. Chin J Grassland, 2008, 30(5): 90-95. |
| [12] |
刘明, 王继华, 王同昌. DNA分子标记技术[J]. 东北林业大学学报, 2003, 31(6): 65-67. Liu M, Wang JH, Wang TC. DNA molecular markers[J]. J Northeast For Univ, 2003, 31(6): 65-67. DOI:10.3969/j.issn.1000-5382.2003.06.023 |
| [13] |
Batsuren E, Zhang X, Song MJ, et al. Density-dependent changes of mating system and family structure in Brandt's voles (Lasiopodomys brandtii)[J]. Ecol Evol, 2022, 12(8): e9199. DOI:10.1002/ece3.9199 |
| [14] |
白晓慧, 蔡超, 郝爽, 等. 大鳞副泥鳅5个野生群体的遗传多样性分析[J]. 基因组学与应用生物学, 2019, 38(7): 2958-2965. Bai XH, Cai C, Hao S, et al. Genetic diversity analysis of five wild Paramisgurnus dabryanus populations[J]. Genomics Appl Biol, 2019, 38(7): 2958-2965. DOI:10.13417/j.gab.038.002958 |
| [15] |
刘蓬勃, 孙琬琬, 王君, 等. 滇西南地区埃及伊蚊种群微卫星标记筛选研究[J]. 中国媒介生物学及控制杂志, 2018, 29(2): 130-133. Liu PB, Sun WW, Wang J, et al. Isolation of microsatellite markers in Aedes aegypti population from southwest Yunnan[J]. Chin J Vector Biol Control, 2018, 29(2): 130-133. DOI:10.11853/j.issn.1003.8280.2018.02.004 |
| [16] |
姜姗, 王兴亚, 王小奇. 基于微卫星标记的中国东北地区灰飞虱遗传变异及种群遗传结构分析[J]. 昆虫学报, 2020, 63(1): 73-84. Jiang S, Wang XY, Wang XQ. Genetic variation and population genetic structure of the small brown planthopper, Laodelphax striatellus (Hemiptera: Delphacidae), in Northeast China based on microsatellite markers[J]. Acta Entomol Sin, 2020, 63(1): 73-84. DOI:10.16380/j.kcxb.2020.01.009 |
| [17] |
康芬芬, 李志红, 杨定, 等. 利用微卫星标记初步分析橘小实蝇4个地理种群的遗传多态性[J]. 昆虫知识, 2006, 43(3): 371-374. Kang FF, Li ZH, Yang D, et al. Microsatellite markers for genetic polymorphism in four geographic populations of the Oriental Fruit Fly, Bactrocera dorsalis[J]. Chin Bull Entomol, 2006, 43(3): 371-374. DOI:10.3969/j.issn.0452-8255.2006.03.023 |
| [18] |
Wang D, Shi DZ. Isolation and characterization of polymorphic microsatellite loci from Brandt's voles (Lasiopodomys brandtii)[J]. Mol Ecol Notes, 2007, 7(4): 671-673. DOI:10.1111/j.1471-8286.2006.01673.x |
| [19] |
牟媛媛. 布氏田鼠婚配制度初探: 基于围栏种群的微卫星研究[D]. 曲阜: 曲阜师范大学, 2019. Mou YY. Mating system of Lasiopodomys brandti: Based on a microsatellite study & the fenced population[D]. Qufu: Qufu Normal University, 2019 (in Chinese) |
| [20] |
唐男男. 增雨对布氏田鼠种群遗传多样性的影响[D]. 曲阜: 曲阜师范大学, 2017. Tang NN. Effect of increased rainfall on the genetic diversity of Brand's vole populations[D]. Qufu: Qufu Normal University, 2017. (in Chinese) |
| [21] |
褚翔宇. 增食对布氏田鼠种群数量及遗传多样性的效应[D]. 曲阜: 曲阜师范大学, 2017. Chu XY. The effect of increased feeding on the population size and genetic diversity of Brandt's vole[D]. Qufu: Qufu Normal University, 2017. (in Chinese) |
| [22] |
魏勇, 胡珂, 陈钰蓝, 等. 白纹伊蚊微卫星标记的筛选及种群遗传研究[J]. 现代预防医学, 2020, 47(7): 1267-1270, 1290. Wei Y, Hu K, Chen YL, et al. Screening of microsatellite markers of Aedes albopictus and their application in the population genetics[J]. Mod Prev Med, 2020, 47(7): 1267-1270, 1290. |
| [23] |
张恒端. 南京市白纹伊蚊种群遗传学研究[D]. 北京: 解放军军事医学科学院, 2015. DOI: 10.7666/d.Y2993915. Zhang HD. Study on population genetics characteristics of Aedes albopictus in Nanjing city[D]. Beijing: Academy of Military Medical Sciences, 2015. DOI: 10.7666/d.Y2993915.(in Chinese) |
| [24] |
Takezaki N, Nei M. Genetic distances and reconstruction of phylogenetic trees from microsatellite DNA[J]. Genetics, 1996, 144(1): 389-399. DOI:10.1093/genetics/144.1.389 |
| [25] |
Andreassen HP, Sundell J, Ecke F, et al. Population cycles and outbreaks of small rodents: Ten essential questions we still need to solve[J]. Oecologia, 2021, 195(3): 601-622. DOI:10.1007/s00442-020-04810-w |
| [26] |
赵庆华. 大仓鼠(Tscherskia triton)种群年际遗传变异与生态适应研究[D]. 曲阜: 曲阜师范大学, 2007. DOI: 10.7666/d.Y1078164. Zhao QH. Interannual genetic variation and ecological adaptation in Tscherskia triton populations[D]. Qufu: Qufu Normal University, 2007. DOI: 10.7666/d.Y1078164.(in Chinese) |
| [27] |
Wang D, Guo YW, Shi DZ. Genetic structure of Brandt's vole (Lasiopodomys brandtii) populations in Inner Mongolia, China, based on microsatellite analysis[J]. Conserv Genet, 2011, 12(3): 659-667. DOI:10.1007/s10592-010-0172-y |
| [28] |
田玉苗, 盛清宇, 袁立成, 等. 长白山北部西伯利亚狍局域种群间的基因流[J]. 野生动物学报, 2020, 41(3): 551-559. Tian YM, Sheng QY, Yuan LC, et al. Gene flow in local populations of Siberian Roe Deer (Capreolus pygargus) in the northern Changbaishan range of China[J]. Chin J Wildlife, 2020, 41(3): 551-559. DOI:10.19711/j.cnki.issn2310-1490.2020.03.001 |
| [29] |
曲若竹, 侯林, 吕红丽, 等. 群体遗传结构中的基因流[J]. 遗传, 2004, 26(3): 377-382. Qu RZ, Hou L, Lyu HL, et al. The gene flow of population genetic structure[J]. Hereditas, 2004, 26(3): 377-382. DOI:10.3321/j.issn:0253-9772.2004.03.022 |