 2023, Vol. 34
2023, Vol. 34扩展功能
文章信息
- 吴洪潘, 刘冬志, 初雯雯, 古丽娜·再努拉, 戚英杰, 初红军
- WU Hong-pan, LIU Dong-zhi, CHU Wen-wen, GULINA Zainula, QI Ying-jie, CHU Hong-jun
- 新疆卡拉麦里山有蹄类野生动物自然保护区大沙鼠巢域微生境选择
- Microhabitat selection of Rhombomys opimus hideout in Mt. Kalamaili Ungulate Nature Reserve, Xinjiang Uygur Autonomous Region, China
- 中国媒介生物学及控制杂志, 2023, 34(1): 70-76
- Chin J Vector Biol & Control, 2023, 34(1): 70-76
- 10.11853/j.issn.1003.8280.2023.01.013
-
文章历史
- 收稿日期: 2022-11-04




2 汕头市濠江区乡村振兴战略发展中心, 广东 汕头 515071;
3 新疆卡拉麦里山有蹄类野生动物自然保护区管理中心, 新疆 乌鲁木齐 830000;
4 新疆环疆绿源环保科技有限公司, 新疆 乌鲁木齐 830019;
5 北京林业大学, 北京 100083;
6 新疆林科院森林生态研究所, 新疆 乌鲁木齐 830002;
7 新疆维吾尔自治区草原总站, 新疆 乌鲁木齐 830049
2 Shantou Haojiang District Rural Revitalization Strategy Development Center, Shantou, Guangdong 515071, China;
3 Management Center, Kalamaili Ungulate Nature Reserve, Urumqi, Xinjiang 830000, China;
4 Huanjiang Lvyuan Environmental Protection Technology Co., Ltd., Urumqi, Xinjiang 830019, China;
5 Beijing Forestry University, Beijing 100083, China;
6 Institute of Forest Ecology, Xinjiang Academy of Forestry Sciences, Urumqi, Xinjiang 830002, China;
7 Grassland Station of Xinjiang Uygur Autonomous Region, Urumqi, Xinjiang 830049, China
大沙鼠(Rhombomys opimus)是啮齿目(Rodentia)沙鼠亚科(Gerbillinae)中最大的类群,是亚洲中部荒漠的典型啮齿动物,是荒漠、半荒漠景观中生物多样性的指示物种之一,主要栖息在丘陵山谷荒漠地区、平原沙土荒漠和粘土荒漠地区,营群居生活[1-3]。因大沙鼠啃食、挖掘巢域周围植物,常造成植物死亡,对荒漠中的梭梭(Haloxylon ammodendron)及其他固沙灌木造成极其严重的危害。大沙鼠国内主要分布在内蒙古、宁夏、甘肃和新疆等省和自治区,国外主要分布在伊朗、阿富汗、俄罗斯、哈萨克斯坦、蒙古国等国家的荒漠地带[4-6]。国内外对大沙鼠的研究主要集中在疫源疫病[7-9]、鼠害[10-11]、寄生虫[12-13]、种群数量[4, 14]、生境选择[15-16]、鼠洞覆盖[6, 17]、分子标记[18]和巢域植被种类等方面[19]。近年的研究发现,大沙鼠是准噶尔盆地鼠疫自然疫源地的鼠疫主要宿主动物,同时还是多种疾病的宿主动物,如皮肤利什曼病、李斯特菌感染、蜱媒回归热等[2-3, 12, 20],但关于其巢域微生境选择的研究和报道较少。
大沙鼠是新疆卡拉麦里山有蹄类野生动物自然保护区(以下简称卡山保护区)内分布范围较广的昼行性鼠类,多种生境类型都有其活动痕迹。在2019年春、夏季及2020年春、夏和秋季,作者调查研究了卡山保护区大沙鼠对巢域的生境选择偏好,分析了其巢域微生境与环境因子之间的关系,结果如下。
1 材料与方法 1.1 自然概况新疆卡山保护区位于准噶尔盆地的东北部,古尔班通古特沙漠西缘,东经88°30′~90°00′,北纬44°36′~46°00′,东西宽117.5 km,南北长147.5 km,面积14 856.48 km2。阿富准铁路、国道G216和新线高速公路从保护区中间穿过,呈半开放性状态[20]。景观主要包括戈壁、沙漠、平原以及山地丘陵等地形地貌。卡山保护区深处亚洲中部腹地,属典型大陆性气候,年平均气温2 ℃,7月平均温度20.5 ℃,1月平均温度-24.3 ℃。年平均降水量低至159.1 mm,年平均蒸发量为2 090 mm。受地理位置、气候和土壤等生态因素的制约,卡山保护区荒漠植被稀少,结构单调,植被覆盖稀疏。植被主要由麻黄科(Ephedra)、柽柳科(Tamaricaeae)和蒺藜科(Zygophyllaceae)的灌木和矮灌木组成。主要建群植物以菊科(Lompositae)、藜科(Ehenopodirm)和柽柳科的种类较多,其次为蒺藜科、禾本科(Gramineae)[21]。常见的灌木有梭梭、木蓼(Atraphaxis frutescens)、沙拐枣(Calligonum mongolicum)、驼绒藜(Ceratoides latens)等[22]。常见的草本植物有假木贼(Anabasis salsa)、芨芨草(Achnatherum splendens)、角果藜(Ceratocarpus arenarius)等。卡山保护区栖息着野放普氏野马(Equus ferus)、蒙古野驴(E. hemionus)、盘羊(Ovis ammon)、鹅喉羚(Gazella subgutturosa)和猞猁(Lynx lynx)等多种国家重点保护野生动物[23-26]。
1.2 研究方法 1.2.1 样方设置及生境因子2019年春、夏季,2020年春、夏和秋季,在卡山保护区调查大沙鼠巢域洞群时,参照李叶等[27]的方法,设置5 m×5 m大样方,四角及中心设置5个1 m×1 m小样方,然后在洞群0.5~1 km以外生境中选择无大沙鼠活动的区域设置对照地样方。记录2种样方的海拔高度、植物种数、植被盖度、植被高度、土壤含水率以及距道路距离等因子,生境因子具体描述如下:
海拔(elevation,E):通过全球定位系统(GPS)记录样方中心点的海拔高度。
坡度(slope,S):使用DQL-4军用罗盘测量5 m×5 m样方中心的坡度值。
植物种数(No. plant species,NPS):计数小样方内的植物种类。
植被盖度(vegetation density,VD):测算5个1 m×1 m小样方内的植被盖度,之后计算其平均值作为大样方的植被盖度。
生物量(biomass,B):剪割5个1 m×1 m小样方内的灌木和草本植物新生部分,120 ℃烘箱恒温烘烤12 h,恒重之后计算其平均值作为大样方的生物量。
灌木种数(number of shrub species,NSS):测定5个1 m×1 m小样方内的灌木种类数,之后计算其平均值作为大样方的灌木种数。
灌木盖度(species density,SD):测算5个1 m×1 m小样方内的灌木盖度,之后计算其平均值作为大样方的灌木盖度数据。
灌木高度(shrub height,SH):测算5个1 m×1 m小样方内的灌木高度,之后计算其平均值作为大样方的灌木高度数据。
草本种数(number of herbs species,NHS):计数5个1 m×1 m小样方内的草本种类,累加作为大样方的草本种数。
草本盖度(herbal density,HD):测算5个1 m×1 m小样方内的草本盖度,之后计算其平均值作为大样方的草本盖度数据。
草本高度(herbal height,HH):测算5个1 m×1 m小样方内的草本高度,之后计算其平均值作为大样方的草本高度数据。
土壤含水率(soil moisture content,SMC):在小样方内用土钻钻取地表 10~30 cm地表土,剔除植物根段和枯落物等杂质,随机取样3次混合,编号带回实验室,置于120 ℃烘箱中烘烤12 h,前后重量的比值作为土壤含水率。
距道路距离(distance to road,DR):测算大样方中心与研究区域最近的便道、乡道和国道的距离。
距水源地距离(distance to water hole,DWH):测算大样方中心与研究区域水源地的距离。
距居民点距离(distance to residential area,DRA):测算大样方中心与研究区域居民点的距离。
距隐蔽物距离(distance to hidden object,DHO):测算大样方中心与研究区域最近隐蔽物(鼠洞、灌木、草丛、沙丘等)的距离。
土壤基质(soil matrix,SM):分为5种,分别为碱土、壤土、黏质土、砂质土、裸岩。
坡位(slope position,SP):分为4种,分别为平滩、坡顶、山腰、坡底。
1.2.2 数据分析在Excel 2019软件中录入野外采集数据,用SPSS 20.0软件进行数据的统计和分析。对数值型生态因子数据分别进行描述性统计分析,确定其各自的均值及标准误。对数据进行单个样本Kolmogorov-Smirnov检验,以判定数据是否符合正态分布。经检验不符合正态分布(P < 0.05)的数据,利用非参数估计中的2个独立样本的Mann-Whitey U检验判定生态因子数据的差异性。经检验符合正态分布(P > 0.05)的数据,利用独立样本t检验判定生态因子数据的差异性。对非数值型生态因子数据进行2个等级变量间的相关分析。检验水准取0.05。
对生境因子数据进行主成分分析(principal component analysis,PCA)。根据样本数据矩阵计算出样本相关矩阵,求出相关矩阵的特征根和特征向量。根据特征根和特征向量求出各主成分及贡献率。通过主成分分析确定在大沙鼠巢域微生境选择上起主要作用的生境因子。
2 结果 2.1 大沙鼠微生境选择的生境因子共调查大沙鼠巢域微生境样方111个,对照样方111个,所获巢域微生境选择相关因子的描述性统计和差异性比较结果见表 1。
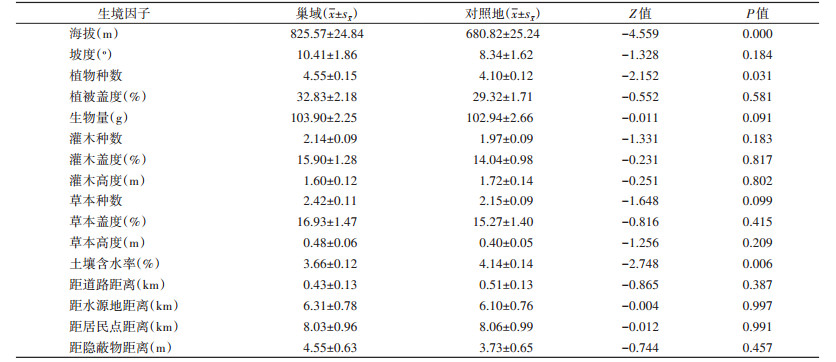
|
在所列16个数值型生境因子中,大沙鼠巢域与对照地2个独立样本的Mann-Whitney U检验表明:大沙鼠巢域与对照地在海拔高度、土壤含水率和植物种数3个生境因子方面的差异均有统计学意义(均P < 0.05)。而其他13个生境因子方面均无统计学意义(均P > 0.05)。
通过2个等级变量间的相关分析,发现大沙鼠巢域微生境与对照地在坡位的选择上,大沙鼠巢域微生境的选择与坡位相关关系极显著,Kendall’s相关系数为0.324,P < 0.001,Spearman相关系数为0.338,P < 0.001。在土壤类型的选择上,大沙鼠巢域微生境与土壤类型相关关系极显著,Kendall’s相关系数为0.370,P < 0.001,Spearman相关系数为0.405,P < 0.001。
2.2 大沙鼠巢域微生境的主成分分析通过主成分分析,得出卡山保护区大沙鼠微生境因子选择的特征值。见表 2。
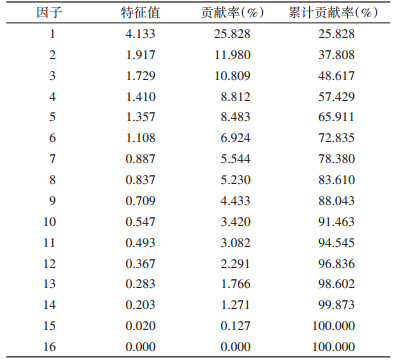
|
主成分分析结果显示,大沙鼠生境选择的相关矩阵前6个主成分的特征值均 > 1,累积贡献率为72.835%。说明前6个因子包含了基本信息,可以较好地反映大沙鼠的微生境特征,因此,选用前6个主成分进行分析,其相应的因子负载见表 3。
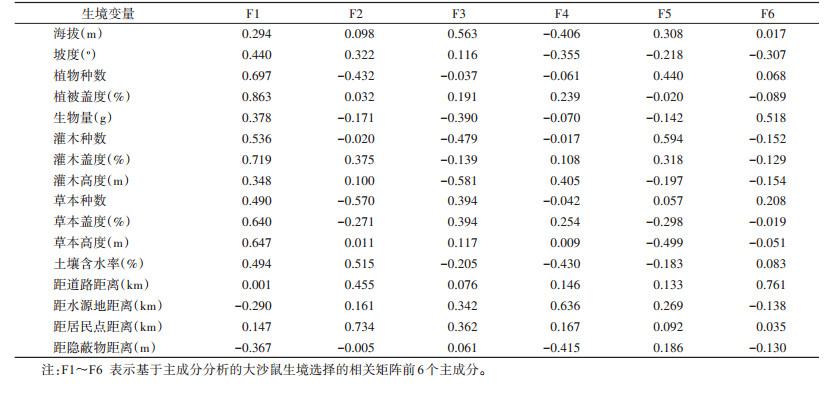
|
由表 2、3可知,第1主成分的贡献率达25.828%,其中绝对值较大的权系数出现在植被盖度(0.863)、灌木盖度(0.719)等生态因子上,显示这些生态因子具有较大的信息荷载量。第2主成分的贡献率为11.980%,其中绝对值较大的权系数出现在距居民点距离(0.734)等生境因子上。第3主成分的贡献率为10.809%,其中绝对值较大的权系数出现在海拔(0.563)和灌木高度(-0.581)等生境因子上。第4主成分的贡献率为8.812%,其中绝对值较大的权系数出现在距水源地距离(0.636)等生境因子上。第5主成分的贡献率为8.483%,其中绝对值较大的权系数出现在灌木种数(0.594)等生境因子上。第6主成分的贡献率为6.924%,其中绝对值较大的权系数出现在距道路距离(0.761)等生境因子上。
3 讨论啮齿类与其所处环境间存在着错综复杂的相互联系,其种群密度受到多方面因素的制约[27]。有关东北鼢鼠(Myospalax psilurus)、布氏田鼠(Microtus brandti)、大沙鼠、高原鼠兔(Ochotona curzoniae)生境选择的研究结果已被用于生态防治,并取得良好的防治效果[15-16, 27-29]。因此研究啮齿动物的微生境选择既有理论意义,又有实践应用价值。
与对照地相比,大沙鼠巢域微生境选择更偏爱海拔高、土壤含水率低、植物种数多、平滩类型,包含砂质土和碱土两种土壤基质的微生境作为巢域。与其他环境因素相比,大沙鼠对海拔高度的选择表现出显著差异,这可能与所处研究区域的地形地貌有关。天敌对捕食场的选择能显著提高捕获率,而啮齿动物对微生境选择则可以躲避天敌,增加存活率[30-31]。天敌猛禽会占据地形高处搜寻猎物,伺机俯冲扑食。李叶等[27]认为高原鼠兔选择微生境倾向于海拔高,坡度陡,植被高度低、密度高,土壤硬度小的高寒草地或高寒荒漠,影响其选择的主要因子是食物和天敌。与高原鼠兔类似,大沙鼠在荒漠环境选择巢域微生境时也会优先选择地势相对开阔且利于奔跑、躲避天敌的平滩地形,及植物种数多的环境获取食物。野外观察发现,卡山保护区中部、北部大沙鼠站立观察警戒周围的情况较多,而卡山保护区南部、西部的大沙鼠站立观察警戒周围的情况少,推测这可能与其反捕食策略有关[32]。保护区北部和中部,受山地丘陵地形遮蔽的影响,猛禽随时会出现,捕食压力大,站立观察能警戒更远范围,提前发现天敌,耗费在警戒等行为的时间多;而南部和西部的沙漠、平滩地形较平缓,视野开阔,无需站立就能发现远处天敌,同时植被条件良好,能就近获取食物和隐蔽,耗费在警戒等行为的时间少。这与徐峰等[33]对大沙鼠行为的研究结果相同。
主成分分析表明影响大沙鼠巢域微生境选择的主要因子依次是植被盖度(0.863)、距道路距离(0.761)、距居民点距离(0.734)、灌木盖度(0.719)。植被因子与食物丰富度密切相关,巢域微生境尺度上对植物种数多、植物盖度高、灌木盖度高等的选择说明大沙鼠偏好在食物丰富度较高的区域筑洞。一方面可获得充足食物,另一方面可减少觅食活动距离,便于隐蔽和减少被捕食风险。而巢域与对照区的植被盖度、种数存在差异,这可能是自然选择的结果。大沙鼠有针对性地选择合适的巢域筑洞,而有蹄类动物会避免在啮齿动物的巢域采食,防止腿部陷入受伤。两者选择的结果,形成植被因子的差异。
巢域的环境因素会对啮齿动物产生不同的影响,对于繁殖力高、营家族生活的大沙鼠,巢域的选择直接影响其繁殖、生存和种群密度。实验表明,卡山保护区内影响大沙鼠巢域选择的第6和第2主成分中,距道路和居民点距离的权系数绝对值较高(0.761、0.734)。这表明在巢域的选择过程中,首先选择在远离道路和居民点的位置建设巢域。回避人类活动的干扰是绝大多数野生动物的选择,但也有研究发现,在人为干扰弱的情况下,卡山保护区的大沙鼠会主动穿越道路[25, 34]。
在调查样地中不论何种植被类型,大沙鼠多选择土壤含水率较低,同时有砂质土和碱土2种土壤基质的环境作为巢域。土壤含水率是大沙鼠巢域微生境选择的重要指标,通过比较,巢域微生境的土壤含水率[(3.66±0.12)%]低于对照组[(4.14±0.14)%],可能在挖掘洞穴时,土壤含水率低,挖掘阻力小。与此相反,崔庆虎等[35]的研究表明,土壤含水量增加导致草本层植被郁闭度和食物资源增加,土壤硬度下降,有利于根田鼠(Alexandromys oeconomus)建造巢穴。荒漠干旱地区的小型啮齿动物普遍具有耐旱的生理特点,能直接从食物中获取所需水分[36],对水的需求不像大型动物那么强烈,而对食物和躲避天敌的要求较高,会投入更多时间与精力。这从大沙鼠巢域微生境与水源地距离差异不显著可以证实(P=0.997)。此外,地势平坦的丘间地也是大沙鼠的主要建巢地形。戴昆等[15]研究表明,古尔班通古特沙漠的大沙鼠主要选择沙丘间地及固定沙丘为筑巢地形。赵天飙等[16]研究大沙鼠对栖息地的选择,发现大沙鼠最适宜生境为白刺、盐爪爪、红柳等植被环绕的半固定沙丘。在荒漠开阔生境,大沙鼠习惯于在沙丘和灌木下等非开放的微生境活动、取食[37],此生境能提供良好隐蔽条件。
巢域点的选择是人为干扰、地形、反天敌捕食和食物量之间权衡的结果。人为干扰被认为是影响巢域选择的主要因素之一。近年,由于矿业和石油工业等退出保护区,原工矿厂区建筑拆解、土地平复并逐步恢复绿植,人类活动的区域只围绕国道216线、阿富准铁路、G216高速公路、国家公益林管护站和喀木斯特附近的居民点,近几年适宜大沙鼠筑巢穴居的范围可能在扩大。
本研究结果显示,大沙鼠倾向于选择海拔高、土壤含水率低的平滩类型,包含砂质土和碱土两种土壤基质,植被盖度高,灌木盖度高,远离居民点和道路等人为干扰的环境作为巢域。研究结果对了解大沙鼠的分布特点和鼠害的防制工作提供了科学依据。因此,建议改造大沙鼠巢域的原生境,改变洞群周围的植被种类和盖度,可起到一定的防控效果。此外,不同季节气候的变化对大沙鼠的巢域微生境及其对微生境资源利用的影响也是值得进一步研究的内容。
志谢 新疆环疆绿源环保科技有限公司为现场工作提供交通工具,王建忠、杨文国、段黎和王勇利同志参加调查和数据收集;卡山保护区管理中心的李文梅、布兰、梁兴凯、艾代、阿成、蒙坎、蒋峰、马自清、陈明勇、文杰等同志提供了许多帮助;北京林业大学周冉博士、唐丽萍博士给予指导和帮助利益冲突 无
| [1] |
Rajabi M, Mansourian A, Pilesjö P, et al. A spatially explicit agent-based simulation model of a reservoir host of cutaneous leishmaniasis, Rhombomys opimus[J]. Ecol Modell, 2018, 370: 33-49. DOI:10.1016/j.ecolmodel.2017.12.004 |
| [2] |
马勇, 王逢桂, 金善科, 等. 新疆北部地区啮齿动物的分类和分布[M]. 北京: 科学出版社, 1987: 28. Ma Y, Wang FG, Jin SK, et al. Glires (rodents and lagomorphs) of northern Xinjiang and their zoogeographical distribution[M]. Beijing: Science Press, 1987: 28. |
| [3] |
王思博, 杨赣源. 新疆啮齿动物志[M]. 乌鲁木齐: 新疆人民出版社, 1983: 39-40. Wang SB, Yang GY. Rodent fauna of Xinjiang[M]. Urumqi: Xinjiang People's Publishing House, 1983: 39-40. |
| [4] |
尹小平, 田延河, 骄娃, 等. 新疆北部边境口岸大沙鼠种群分布特征及活动规律调查[J]. 中国媒介生物学及控制杂志, 2015, 26(2): 196-199. Yin XP, Tian YH, Jiao W, et al. The distribution characteristics and activity patterns of the Rhombomys opimus population in northern Xinjiang borders[J]. Chin J Vector Biol Control, 2015, 26(2): 196-199. DOI:10.11853/j.issn.1003.4692.2015.02.024 |
| [5] |
Laperrière V, Brugger K, Rubel F. Cross-scale modeling of a vector-borne disease, from the individual to the metapopulation: The seasonal dynamics of sylvatic plague in Kazakhstan[J]. Ecol Modell, 2016, 342: 34-48. DOI:10.1016/j.ecolmodel.2016.09.023 |
| [6] |
刘江, 徐先英, 张荣娟, 等. 人工梭梭林大沙鼠(Rhombomys opimus)土丘斑块与鼠洞塌陷斑块破碎度及点格局[J]. 中国沙漠, 2018, 38(3): 610-618. Liu J, Xu XY, Zhang RJ, et al. Fragmentation and point pattern of mound patches and hole collapse patches of Rhombomys opimus in the artificial Haloxylon ammodendron plantations[J]. J Desert Res, 2018, 38(3): 610-618. DOI:10.7522/j.issn.1000-694X.2017.00036 |
| [7] |
Foroutan M, Khademvatan S, Majidiani H, et al. Prevalence of Leishmania species in rodents: A systematic review and meta-analysis in Iran[J]. Acta Trop, 2017, 172: 164-172. DOI:10.1016/j.actatropica.2017.04.022 |
| [8] |
Shiravand B, Tafti AAD, Hanafi-Bojd AA, et al. Modeling spatial risk of zoonotic cutaneous leishmaniasis in Central Iran[J]. Acta Trop, 2018, 185: 327-335. DOI:10.1016/j.actatropica.2018.06.015 |
| [9] |
Nilsson P, Solbakken MH, Schmid BV, et al. The genome of the great gerbil reveals species-specific duplication of an MHCII gene[J]. Genome Biol Evol, 2020, 12(2): 3832-3849. DOI:10.1093/gbe/evaa008 |
| [10] |
杨永刚, 陈梦, 刘忠军, 等. 三项调查指标与大沙鼠对林木危害率的相关性分析[J]. 中国森林病虫, 2014, 33(1): 11-13. Yang YG, Chen M, Liu ZJ, et al. Correlation analysis between three investigative indexes and damage rate of Rhombomys opimus to forest[J]. For Pest Dis, 2014, 33(1): 11-13. DOI:10.3969/j.issn.1671-0886.2014.01.004 |
| [11] |
Xu WX, Liu W, Yang WK, et al. Impact of great gerbils (Rhombomys opimus) on desert plant communities[J]. J Arid Land, 2015, 7(6): 852-859. DOI:10.1007/s40333-015-0084-2 |
| [12] |
Zhang YJ, Dai X, Wang QG, et al. Transmission efficiency of the plague pathogen (Y. pestis) by the flea, Xenopsylla skrjabini, to mice and great gerbils[J]. Parasit Vectors, 2015, 8: 256. DOI:10.1186/s13071-015-0852-z |
| [13] |
Gholipoury M, Rezai HR, Namroodi S, et al. Zoonotic and non-zoonotic parasites of wild rodents in Turkman Sahra, Northeastern Iran[J]. Iran J Parasitol, 2016, 11(3): 350-357. |
| [14] |
盛兆湖, 陈梦, 刘忠军, 等. 新疆昌吉州荒漠林区鼠类群落结构与梭梭被害关系的研究[J]. 中国森林病虫, 2014, 33(2): 4-7. Sheng ZH, Chen M, Liu ZJ, et al. Relationship between the community structure of rodents and damage on Haloxylon ammodendrom in the desert forest area in Changji, Xinjiang[J]. For Pest Dis, 2014, 33(2): 4-7. DOI:10.3969/j.issn.1671-0886.2014.02.002 |
| [15] |
戴昆, 姚军, 高行宜. 大沙鼠的巢域选择[J]. 干旱区研究, 1999, 16(4): 5-9. Dai K, Yao J, Gao XY. Home range choose of great gerbil (Rhombomys opimus)[J]. Arid Zone Res, 1999, 16(4): 5-9. DOI:10.13866/j.azr.1999.04.002 |
| [16] |
赵天飙, 张忠兵, 李新民, 等. 大沙鼠对栖息地的选择[J]. 动物学杂志, 2000, 35(1): 40-43. Zhao TB, Zhang ZB, Li XM, et al. Habitat selection of Rhombomys opimus[J]. Chin J Zool, 2000, 35(1): 40-43. DOI:10.3969/j.issn.0250-3263.2000.01.017 |
| [17] |
马涛, 郑江华, 温阿敏, 等. 基于UAV低空遥感的荒漠林大沙鼠洞群覆盖率及分布特征研究: 以新疆古尔班通古特沙漠南缘局部为例[J]. 生态学报, 2018, 38(3): 953-963. Ma T, Zheng JH, Wen AM, et al. Group coverage of burrow entrances and distribution characteristics of desert forest-dwelling Rhombomys opimus based on unmanned aerial vehicle (UAV) low-altitude remote sensing: A case study at the southern margin of the Gurbantunggut Desert in Xinjiang[J]. Acta Ecol Sin, 2018, 38(3): 953-963. DOI:10.5846/stxb201612142571 |
| [18] |
Hosseinzadeh-Colagar A, Haghighatnia MJ, Amiri Z, et al. Microsatellite (SSR) amplification by PCR usually led to polymorphic bands: Evidence which shows replication slippage occurs in extend or nascent DNA strands[J]. Mol Biol Res Commun, 2016, 5(3): 167-174. |
| [19] |
杨维康, 乔建芳, 蒋慧萍, 等. 大沙鼠掘洞对准噶尔荒漠植物群落的小尺度影响[J]. 干旱区地理, 2006, 29(2): 219-224. Yang WK, Qiao JF, Jiang HP, et al. Impact of burrows of the great gerbil on small-scale plant community in Junggar Desert, China[J]. Arid Land Geogr, 2006, 29(2): 219-224. DOI:10.13826/j.cnki.cn65-1103/x.2006.02.009 |
| [20] |
Rajabi M, Mansourian A, Pilesjö P, et al. A spatially explicit agent-based simulation model of a reservoir host of cutaneous leishmaniasis, Rhombomys opimus[J]. Ecol Modell, 2018, 370: 33-49. DOI:10.1016/j.ecolmodel.2017.12.004 |
| [21] |
陈晨, 邵长亮, 葛炎, 等. 卡拉麦里山有蹄类野生动物自然保护区蒙古野驴生境格局动态及其成因分析[J]. 生态学报, 2021, 41(5): 2056-2066. Chen C, Shao CL, Ge Y, et al. Habitat pattern dynamics and cause analysis of Equus hemionus in Kalamaili Mountain Ungulate Nature Reserve, Xinjiang[J]. Acta Ecol Sin, 2021, 41(5): 2056-2066. DOI:10.5846/stxb202004010773 |
| [22] |
Xia CJ, Cao J, Zhang HF, et al. Reintroduction of Przewalski's horse (Equus ferus przewalskii) in Xinjiang, China: The status and experience[J]. Biol Conserv, 2014, 177: 142-147. DOI:10.1016/j.biocon.2014.06.021 |
| [23] |
初红军, 蒋志刚, 葛炎, 等. 卡拉麦里山有蹄类自然保护区蒙古野驴和鹅喉羚种群密度和数量[J]. 生物多样性, 2009, 17(4): 414-422. Chu HJ, Jiang ZG, Ge Y, et al. Population densities and number of Khulan and Goitred gazelle in Mt. Kalamaili Ungulate Nature Reserve[J]. Biodiv Sci, 2009, 17(4): 414-422. DOI:10.3724/SP.J.1003.2009.09001 |
| [24] |
Zhang YJ, Cao QS, Rubenstein DI, et al. Water use patterns of sympatric Przewalski's horse and Khulan: Interspecific comparison reveals niche differences[J]. PLoS One, 2015, 10(7): e0132094. DOI:10.1371/journal.pone.0132094 |
| [25] |
Ji SN, Jiang ZG, Li LL, et al. Impact of different road types on small mammals in Mt. Kalamaili Nature Reserve[J]. Transp Res D Transp Environ, 2017, 50: 223-233. DOI:10.1016/j.trd.2016.11.006 |
| [26] |
国家林业和草原局农业农村部公告(2021年第3号): 国家重点保护野生动物名录[EB/OL]. (2021-02-05)[2022-11-04]. http://www.forestry.gov.cn/main/3457/20210205/122612568723707.html. Announcement of the National Forestry and Grassland Administration and the Ministry of Agriculture and Rural Affairs of the People's Republic of China (No. 3 of 2021): List of national key protected wild animals[EB/OL]. (2021-02-05)[2022-11-04]. http://www.forestry.gov.cn/main/3457/20210205/122612568723707.html. |
| [27] |
李叶, 王振宇, 张翔, 等. 阿尔金山自然保护区高原鼠兔夏季微生境选择的主导因子分析[J]. 中国媒介生物学及控制杂志, 2014, 25(1): 28-31. Li Y, Wang ZY, Zhang X, et al. Analysis of dominant factors affecting microhabitat selection of plateau pika (Ochotona curzoniae) during summer in Altun Mountain National Nature Reserve, Xinjiang Uygur Autonomous Region, China[J]. Chin J Vector Biol Control, 2014, 25(1): 28-31. DOI:10.11853/j.issn.1003.4692.2014.01.008 |
| [28] |
钟文勤, 周庆强, 孙崇潞. 布氏田鼠的生境选择与植被条件[M]//中国科学院内蒙古草原生态系统定位站. 草原生态系统研究. 第1集. 北京: 科学出版社, 1983: 107-111. Zhong WQ, Zhou QQ, Sun CL. Habitat selection and vegetation conditions of Microtus brandti[M]//Inner Mongolia Grassland Ecosystem Research Station, Chinese Academy of Sceiences. Study on grassland ecosystem. Beijing: Science Press, 1983: 107-111. (in Chinese) |
| [29] |
刘仁华, 刘炳友, 赵秀成, 等. 林区鼢鼠鼠害的主要特征及其生态控制对策[J]. 兽类学报, 1997, 17(4): 272-278. Liu RH, Liu BY, Zhao XC, et al. The basic characteristics of the zokor pests and the ecological strategies of controlling in forest area[J]. Acta Theriol Sin, 1997, 17(4): 272-278. DOI:10.16829/j.slxb.1997.04.006 |
| [30] |
王振宇, 李叶, 张翔, 等. 高原兔夏季卧栖地生境利用关键因子分析[J]. 四川动物, 2015, 34(1): 47-52. Wang ZY, Li Y, Zhang X, et al. Analysis of dominant factors affecting resting sites use of Lepus oiostolus in summer[J]. Sichuan J Zool, 2015, 34(1): 47-52. DOI:10.3969/j.Issn.1000-7083.2015.01.007 |
| [31] |
Kajtoch Ł, Żmihorski M, Wieczorek P. Habitat displacement effect between two competing owl species in fragmented forests[J]. Popul Ecol, 2015, 57(3): 517-527. DOI:10.1007/s10144-015-0497-y |
| [32] |
Halle S. Diel pattern of predation risk in microtine rodents[J]. Oikos, 1993, 68(3): 510-518. DOI:10.2307/3544919 |
| [33] |
徐峰, 刘伟, 乔洪海, 等. 大沙鼠的警戒行为与采食及其它行为之间的关系[J]. 干旱区地理, 2013, 36(1): 118-124. Xu F, Liu W, Qiao HH, et al. Relationships between vigilance, foraging and other behaviors of the great gerbil Rhombomys opimus[J]. Arid Land Geogr, 2013, 36(1): 118-124. DOI:10.13826/j.cnki.cn65-1103/x.2013.01.002 |
| [34] |
汪沐阳, 吉晟男, 邵长亮, 等. 不同类型道路对卡拉麦里山有蹄类野生动物自然保护区大沙鼠分布的影响[J]. 四川动物, 2021, 40(1): 1-7. Wang MY, Ji SN, Shao CL, et al. Effects of road type on the distribution of great gerbil in the Kalamaili Mountain Ungulate Nature Reserve, China[J]. Sichuan J Zool, 2021, 40(1): 1-7. DOI:10.11984/j.issn.1000-7083.20200316 |
| [35] |
崔庆虎, 蒋志刚, 连新明, 等. 根田鼠栖息地选择的影响因素[J]. 兽类学报, 2005, 25(1): 45-51. Cui QH, Jiang ZG, Lian XM, et al. Factors influencing habitat selection of root voles (Microtus oeconomus)[J]. Acta Theriol Sin, 2005, 25(1): 45-51. DOI:10.16829/j.slxb.2005.01.008 |
| [36] |
王定国. 额济纳旗和肃北马鬃山北部边境地区啮齿动物调查[J]. 动物学杂志, 1988, 23(6): 21-24. Wang DG. Investigation on rodents in the border areas of Ejine banner and northern Mazong Mountain in Subei[J]. Chin J Zool, 1988, 23(6): 21-24. DOI:10.13859/j.cjz.1988.06.008 |
| [37] |
周立志, 马勇, 李迪强. 大沙鼠在中国的地理分布[J]. 动物学报, 2000, 46(2): 130-137. Zhou LZ, Ma Y, Li DQ. Distribution of great gerbil (Rhombomys opimus) in China[J]. Acta Zool Sin, 2000, 46(2): 130-137. DOI:10.3969/j.issn.1674-5507.2000.02.002 |