 2023, Vol. 34
2023, Vol. 34扩展功能
文章信息
- 张璐, 韦晓慧, 常楠, 李泓运, 司晓燕, 白雪峰, 王君, 刘起勇
- ZHANG Lu, WEI Xiao-hui, CHANG Nan, LI Hong-yun, SI Xiao-yan, BAI Xue-feng, WANG Jun, LIU Qi-yong
- 内蒙古锡林郭勒盟2012-2021年鼠类寄生蚤构成和蚤指数时空分布特征分析
- Composition of parasitic fleas on rodents and spatiotemporal distribution of flea index in Xilin Gol League, Inner Mongolia Autonomous Region, China, 2012-2021
- 中国媒介生物学及控制杂志, 2023, 34(1): 82-90
- Chin J Vector Biol & Control, 2023, 34(1): 82-90
- 10.11853/j.issn.1003.8280.2023.01.015
-
文章历史
- 收稿日期: 2022-10-18




2 南京医科大学, 江苏 南京 210000;
3 山东第一医科大学(山东省医学 科学院), 山东 济南 271016;
4 内蒙古自治区综合疾病预防控制中心, 内蒙古 呼和浩特 010020;
5 锡林郭勒盟疾病预防控制中心, 内蒙古 锡林浩特 026099
2 School of Public Health, Nanjing Medical University, Nanjing, Jiangsu 210000, China;
3 School of Public Health, Shandong First Medical University & Shandong Academy of Medical Sciences, Ji'nan, Shandong 271016, China;
4 Inner Mongolia Comprehensive Disease Control and Prevention Center, Hohhot, Inner Mongolia 010020, China;
5 Xilin Gol League Center for Disease Control and Prevention, Xilinhot, Inner Mongolia 026099, China
鼠疫是危害人类最严重的烈性传染病之一,其传染性强,传播速度快,致死率高,在我国《传染病防治法》中被列为甲类传染病[1],是我国重要媒介生物传染病之一[2]。锡林郭勒盟位于内蒙古自治区中部,地域辽阔,自然地理景观多样,其所辖12个旗(县)均为鼠疫自然疫源地,其中包含了内蒙古高原长爪沙鼠鼠疫自然疫源地、松辽平原察哈尔丘陵达乌尔黄鼠鼠疫自然疫源地和锡林郭勒高原布氏田鼠鼠疫自然疫源地3种类型疫源地[3]。近年来,锡林郭勒盟鼠疫自然疫源地较为活跃,2012-2021年间全盟共报告3例鼠疫病例;2017-2020年连续4年从鼠体寄生蚤同形客蚤指名亚种(Xenopsylla conformis conformis)和秃病蚤蒙冀亚种(Nosopsyllus laeviceps kuzenkovi)体内分离出鼠疫耶尔森菌(Yersinia pestis,鼠疫菌)。蚤类是鼠疫传播的重要媒介生物,并且在鼠疫自然疫源地延续存在方面起着不可忽视的作用,长期调查和监测锡林郭勒盟鼠体寄生蚤的种类构成和数量分布状况,为深入研究其动态变化规律和有效预防、控制鼠疫疫情发生和流行具有重要意义。
1 材料与方法 1.1 资料来源监测数据来源于中国疾病预防控制信息系统鼠疫防治管理信息系统,包含2012-2021年锡林郭勒盟12个旗(县)(图 1)的鼠体蚤监测数据。

|
| 图 1 内蒙古自治区及锡林郭勒盟鼠疫自然疫源地分布及研究样地 Figure 1 Distribution of study plots and natural plague foci in Xilin Gol League, Inner Mongolia Autonomous Region |
| |
根据《全国鼠疫监测方案》(2005)要求进行调查。鼠体蚤由捕获活体鼠检蚤获取,并对其进行分类鉴定。
1.3 指标定义及计算选用鼠体染蚤率和蚤指数描述鼠体染蚤情况。选用Shannon⁃Wienner多样性指数(H')、Margalef丰富度指数(M)、Pielou均匀度指数(J)和Simpson优势度指数(D)[4]对不同鼠种所携带寄生蚤的群落结构多样性进行分析。

|
式中,S为某鼠种携带寄生蚤物种数,Pi为某鼠种携带第i种蚤个体数(Ni)与蚤总数(N)的比值。
1.4 统计学分析使用Excel 2019软件录入数据,建立数据库并对数据进行整理汇总。使用SPSS 19.0软件对不同鼠种鼠体蚤指数进行统计学分析。采用Kruskal-Wallis H秩和检验[5]进行多组比较。使用R 4.1.3软件,通过Mann-Kenddall检验[6]对10年来不同鼠种鼠体蚤指数的变化趋势进行分析,以P < 0.05为差异有统计学意义。使用PAST 2.17软件计算宿主动物体表寄生蚤群落结构多样性。使用ArcGIS 10.5软件绘制不同鼠种鼠体蚤指数空间分布图。
2 结果 2.1 不同鼠种染蚤基本情况2012-2021年在锡林郭勒盟共捕获鼠4科13属18种,共计50 551只,其中带蚤鼠19 958只,平均染蚤率为39.48%;共捕获蚤类5科14属35种、亚种,共计83 147匹,总蚤指数为1.64;长爪沙鼠(Meriones unguiculatus)(46.28%)、达乌尔黄鼠(Spermophilus dauricus)(27.67%)以及布氏田鼠(Microtus brandti)(16.70%)为当地的优势鼠种。其中,达乌尔黄鼠的染蚤率和蚤指数最高,分别为61.51%和3.43,其次为布氏田鼠(34.17%,0.86)和长爪沙鼠(27.87%,0.76)(表 1)。长爪沙鼠共携带蚤≥29种(亚种),主要为同形客蚤指名亚种和秃病蚤蒙冀亚种,分别占32.31%(5 754/17 809)和30.42%(5 417/17 809);达乌尔黄鼠共携带蚤≥21种(亚种),主要为方形黄鼠蚤蒙古亚种(Citellophilus tesquorum mongolicus),占78.12%(37 498/48 002);布氏田鼠共携带蚤≥19种(亚种),主要为近代新蚤东方亚种(Neopsylla pleskei orientalis)和光亮额蚤(Frontopsylla luculenta),分别占52.94%(3 865/7 301)和27.34%(1 996/7 301)。见表 2。
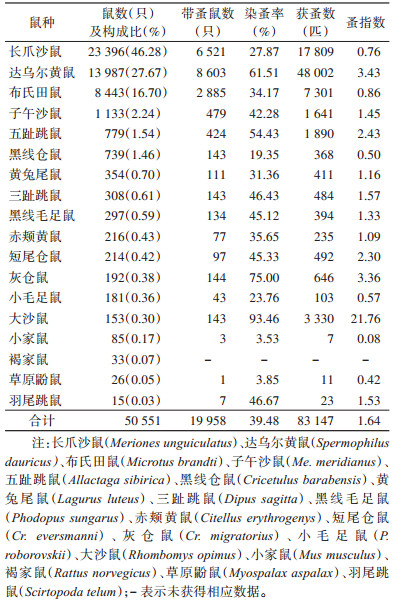
|
长爪沙鼠寄生蚤的D为0.781,H'为1.909,M为2.861,J为0.567;达乌尔黄鼠鼠体寄生蚤的上述4种指数依次分别为0.374、0.827、1.855和0.272;布氏田鼠鼠体寄生蚤的上述4种指数依次分别为0.632、1.325、2.023和0.450(表 2、图 2)。综合各指数结果,长爪沙鼠鼠体寄生蚤的物种多样性高于布氏田鼠和达乌尔黄鼠。

|

|
| 注:D Simpson优势度指数;H' Shannon⁃Wienner多样性指数;M Margalef丰富度指数;J Pielou均匀度指数。 图 2 内蒙古自治区锡林郭勒盟优势鼠种体表寄生蚤的物种多样性 Figure 2 Diversity of ectoparasitic fleas on main rodent species in Xilin Gol League, Inner Mongolia Autonomous Region |
| |
选取混合鼠体蚤指数以及3种优势鼠种鼠体蚤指数进行分析,结果(图 3)显示,2012-2021年,锡林郭勒盟混合鼠体蚤指数(S=-25.000,P=0.032)和达乌尔黄鼠鼠体蚤指数(S=-23.000,P=0.049)整体呈下降趋势。长爪沙鼠(S=-19.000,P=0.107)和布氏田鼠(S=-15.000,P=0.211)的鼠体蚤指数未发现变化趋势。长爪沙鼠、达乌尔黄鼠和布氏田鼠鼠体蚤指数分布不全相同(H=19.355,P < 0.001)。两两比较结果显示,达乌尔黄鼠鼠体蚤指数高于布氏田鼠(H=15.000,P < 0.001)和长爪沙鼠(H=-15.000,P < 0.001),布氏田鼠和长爪沙鼠鼠体蚤指数的差异无统计学意义(H < 0.001,P=1.000)。

|
| 图 3 2012-2021年内蒙古自治区锡林郭勒盟混合鼠体和主要鼠种鼠体蚤指数分布 Figure 3 Distribution of mixed flea index and main rodent flea index in Xilin Gol League, Inner Mongolia Autonomous Region, 2012-2021 |
| |
由于不同鼠种监测的重点时间不同,因此缺乏个别月份的监测数据。混合鼠体蚤指数在7月出现高峰(4.47);长爪沙鼠鼠体蚤指数在3月(1.04)、4月(0.96)较高;达乌尔黄鼠鼠体蚤指数在7月(5.55)、8月(5.68)较高,11月(11.00)最高;布氏田鼠鼠体蚤指数在3月(1.88)较高。见图 4。

|
| 注:1、2和12月无长爪沙鼠和达乌尔黄鼠监测数据;1、2和7月无布氏田鼠监测数据。 图 4 2012-2021年内蒙古自治区锡林郭勒盟混合鼠体及主要鼠种鼠体蚤指数月分布 Figure 4 Monthly distribution of mixed flea index and main rodent flea index in Xilin Gol League, Inner Mongolia Autonomous Region, 2012-2021 |
| |
长爪沙鼠鼠疫自然疫源地监测点包括二连浩特市、苏尼特左旗、苏尼特右旗、太仆寺旗、镶黄旗和正镶白旗,如图 5所示,长爪沙鼠蚤指数较高的地区主要分布在锡林郭勒盟西部,整体分布差异有统计学意义(H=52.579,P < 0.001)。蚤指数最高的是苏尼特左旗和镶黄旗,其与西乌珠穆沁旗、正镶白旗以及正蓝旗差异有统计学意义(均P < 0.05),同时苏尼特左旗高于阿巴嘎旗和苏尼特右旗(均P < 0.05),其他监测点经两两比较差异均无统计学意义(均P > 0.05)。

|
| 注:A 2012-2021年总蚤指数;B~K分别为2012-2021年各年蚤指数。 图 5 2012-2021年内蒙古自治区锡林郭勒盟长爪沙鼠鼠体蚤指数空间分布 Figure 5 Spatial distribution of Meriones unguiculatus flea index in Xilin Gol League, Inner Mongolia Autonomous Region, 2012-2021 |
| |
达乌尔黄鼠鼠疫自然疫源地监测点包括镶黄旗、正镶白旗、正蓝旗和多伦县,但全盟12个监测点10年间均有达乌尔黄鼠监测数据,总体分布差异有统计学意义(H=73.253,P < 0.001)。西乌珠穆沁旗达乌尔黄鼠鼠体蚤指数最高,高于苏尼特左旗、太仆寺旗、二连浩特市、东乌珠穆沁旗以及正蓝旗(均P < 0.05),正镶白旗高于太仆寺旗、二连浩特市、东乌珠穆沁旗以及正蓝旗(均P < 0.05),锡林浩特市高于二连浩特市、东乌珠穆沁旗以及正蓝旗(均P < 0.05),阿巴嘎旗、苏尼特左旗和镶黄旗均高于正蓝旗(均P < 0.05)。结果均显示西乌珠穆沁旗、锡林浩特市以及阿巴嘎旗等布氏田鼠鼠疫自然疫源地的达乌尔黄鼠鼠体蚤指数较高。见图 6。

|
| 注:A 2012-2021年总蚤指数;B~K分别为2012-2021年各年蚤指数。 图 6 2012-2021年内蒙古自治区锡林郭勒盟达乌尔黄鼠鼠体蚤指数空间分布 Figure 6 Spatial distribution of Spermophilus dauricus flea index of in Xilin Gol League, Inner Mongolia Autonomous Region, 2012-2021 |
| |
锡林郭勒高原布氏田鼠鼠疫自然疫源地监测点包括锡林浩特市、阿巴嘎旗、东乌珠穆沁旗和西乌珠穆沁旗,总体上发现,阿巴嘎旗和西乌珠穆沁旗的蚤指数较高,但4个监测点布氏田鼠鼠体蚤指数的差异无统计学意义(H=5.864,P=0.118)。见图 7。

|
| 注:A 2012-2021年总蚤指数;B~K分别为2012-2021年各年蚤指数。 图 7 2012-2021年内蒙古自治区锡林郭勒盟布氏田鼠鼠体蚤指数空间分布 Figure 7 Spatial distribution of Microtus brandti flea index of in Xilin Gol League from, Inner Mongolia Autonomous Region, 2012-2021 |
| |
蚤类是鼠疫传播过程中最重要的媒介生物。蚤指数是指捕捉一定数量的宿主动物,经统计所检出蚤类在某种宿主动物体上的平均染蚤数。鼠体蚤指数对于鼠间鼠疫流行有着重要影响[7]。2012-2021年间,锡林郭勒盟共监测到18种鼠以及其携带的≥35种体表寄生蚤,种类繁多。长爪沙鼠鼠疫疫源地近年来动物间疫情时有发生,主要宿主长爪沙鼠和媒介蚤呈现数量多、分布广的特点[8]。长爪沙鼠体表所携带的主要蚤种同形客蚤指名亚种和秃病蚤蒙冀亚种曾检出鼠疫菌[9],但其他媒介蚤如二齿新蚤[10]和簇鬃客蚤[11]也不能忽视,应加强监测工作。达乌尔黄鼠在3种优势鼠种中的染蚤率和蚤指数最高,以往研究中,达乌尔黄鼠鼠疫自然疫源地主要寄生蚤为方形黄鼠蚤松江亚种[12],但本研究发现,锡林郭勒盟达乌尔黄鼠主要携带的寄生蚤为方形黄鼠蚤蒙古亚种,可能是由于不同疫区的生态环境不同导致。范蒙光等[13]研究发现,近年来达乌尔黄鼠携带的阿巴盖新蚤数量超过原优势蚤种二齿新蚤,本文研究结果与其一致。2012-2021年间,锡林郭勒盟达乌尔黄鼠鼠体蚤指数呈现波动下降趋势,但仍高于兴安盟、通辽市和赤峰市等其他达乌尔黄鼠鼠疫自然疫源地[8],说明鼠疫疫情的不确定性和远距离传播的危险性仍然存在[14]。布氏田鼠鼠疫疫源地自2006年后,一直处于静息状态[15]。本研究分析发现布氏田鼠所携带的主要蚤种为近代新蚤东方亚种和光亮额蚤,而原主要媒介蚤原双蚤田野亚种[16-17]数量降低,这可能与该疫源地部分地区长爪沙鼠数量增多,其携带的近代新蚤东方亚种存在宿主转换有关。
群落多样性与丰富度、均匀度及多样性指数有关,是反映各物种在群落内分布均匀程度及多样性高低的定量指标,优势度指数用于评价优势种在群落内所处的地位和作用[18-19]。Shannon⁃Wienner多样性指数反映不同鼠种所携带寄生蚤的多样性,其越大,表明该鼠种携带的寄生多样性越高;Margalef丰富度指数主要反映的是宿主动物所携带的寄生蚤群落中物种的丰富度,该指数越高,表明寄生蚤种类越丰富;Pielou均匀度指数反映了宿主动物鼠体蚤各个物种的个体数目分配的均匀程度,每个种类的个体数越是接近,群落的均匀度就越高;而Simpson优势度指数主要反映的是宿主动物携带寄生蚤群落中的优势程度,该值越大,说明优势物种越少,多样性越高[20]。本文统计结果显示,长爪沙鼠体表寄生蚤的多样性较高,4个指数均高于布氏田鼠和达乌尔黄鼠,这可能是长爪沙鼠携带并传播鼠疫菌风险较大的原因之一[21]。布氏田鼠携带体表寄生蚤多样性介于长爪沙鼠和达乌尔黄鼠之间,位于第2位。达乌尔黄鼠携带的优势蚤种方形黄鼠蚤蒙古亚种的数量占比高达78.12%,导致其群落结构多样性较低。龚正达等[22]的研究表明,蚤类物种多样性与地理和环境因素相关。因此,锡林郭勒盟3种不同类型的鼠疫疫源地蚤类群落和物种多样性及其与地理环境因素之间关系,值得进一步研究。
已有研究表明,锡林浩特市布氏田鼠鼠疫自然疫源地已演变成与长爪沙鼠鼠疫并存的2种菌型双宿主鼠疫自然疫源地[23]。本文对2012-2021年10年间锡林郭勒盟12个旗(县)鼠体蚤指数分析发现,布氏田鼠鼠疫自然疫源地东乌珠穆沁旗和锡林浩特市与长爪沙鼠鼠疫自然疫源地苏尼特左旗和镶黄旗的长爪沙鼠鼠体蚤指数差异无统计学意义,进一步印证了布氏田鼠鼠疫自然疫源地存在长爪沙鼠鼠疫流行的现象,其原因可能是过度放牧导致草地退化,甚至出现沙化,宜于长爪沙鼠生存[24]。达乌尔黄鼠在全盟各旗(县)均有分布,但布氏田鼠鼠疫自然疫源地的达乌尔黄鼠鼠体蚤指数处于较高水平,以西乌珠穆沁旗最高,其次为锡林浩特市以及阿巴嘎旗,提示该地区除关注原有宿主动物外,应加强对达乌尔黄鼠及其鼠体蚤监测工作。另外,达乌尔黄鼠鼠疫自然疫源地与长爪沙鼠鼠疫自然疫源地在地理分布上交叉重叠[15]。达乌尔黄鼠鼠疫自然疫源地也曾出现过长爪沙鼠鼠疫的流行,如2020年正蓝旗长爪沙鼠检岀了鼠疫菌[8]。因此,上述地区在做好达乌尔黄鼠鼠疫监测的同时,也不能忽视长爪沙鼠鼠疫疫情的监测工作。
综上所述,2012-2021年这10年来锡林郭勒盟主要鼠种的鼠体蚤指数基本处于较平稳状态,但其3种类型鼠疫自然疫源地交叉重叠,长爪沙鼠携带寄生蚤多样性高,传播鼠疫菌风险较大。各监测点应做好动物间鼠疫综合监测工作,警惕各疫源地动物间疫情复燃并波及人间。
利益冲突 无
| [1] |
杨子京, 尹家祥. 我国蚤传自然疫源性疾病流行现状[J]. 包头医学院学报, 2015, 31(2): 143-144. Yang ZJ, Yin JX. Epidemic status of flea borne natural epidemic diseases in China[J]. J Baotou Med Coll, 2015, 31(2): 143-144. DOI:10.16833/j.cnki.jbmc.2015.02.094 |
| [2] |
刘起勇. 2005-2020年我国媒介生物传染病报告病例: 流行趋势、防控挑战及应对策略[J]. 中国媒介生物学及控制杂志, 2022, 33(1): 1-7. Liu QY. Reported cases of vector-borne diseases in China, 2005-2020:Epidemic trend, challenges in prevention and control, and related coping strategies[J]. Chin J Vector Biol Control, 2022, 33(1): 1-7. DOI:10.11853/j.issn.1003.8280.2022.01.001 |
| [3] |
方喜业, 杨瑞馥, 刘起勇, 等. 中国鼠疫自然疫源地分型研究Ⅱ. 鼠疫自然疫源地分型方法研究[J]. 中华流行病学杂志, 2012, 33(2): 234-238. Fang XY, Yang RF, Liu QY, et al. A novel method for typing natural plague foci in China Ⅱ. Research on the typing methods for natural plague foci[J]. Chin J Epidemiol, 2012, 33(2): 234-238. DOI:10.3760/cma.j.issn.0254-6450.2012.02.024 |
| [4] |
Zhao JJ, Gong L, Chen X. Relationship between ecological stoichiometry and plant community diversity in the upper reaches of Tarim River, northwestern China[J]. J Arid Land, 2020, 12(2): 227-238. DOI:10.1007/s40333-019-0020-y |
| [5] |
孙振球. 医学统计学[M]. 3版. 北京: 人民卫生出版社, 2002: 142-145. Sun ZQ. Medical statistics[M]. 3rd ed. Beijing: People's Medical Publishing House, 2002: 142-145. |
| [6] |
Kendall MG. Rank correlation methods[M]. London: Charles Griffin, 1975: 1-202.
|
| [7] |
刘振才, 周晓磊, 张博宇, 等. 动物鼠疫预测模型及预警指标的建立[J]. 中国地方病防治杂志, 2015, 30(1): 1-3. Liu ZC, Zhou XL, Zhang BY, et al. Establishment of predictive model and warning index of animal plague[J]. Chin J Ctrl Endem Dis, 2015, 30(1): 1-3. |
| [8] |
胡艳红, 王姝懿, 李建云, 等. 内蒙古自治区2020年鼠疫监测调查与分析[J]. 中国媒介生物学及控制杂志, 2022, 33(3): 418-425. Hu YH, Wang SY, Li JY, et al. Analysis of plague surveillance in Inner Mongolia Autonomous Region, China, 2020[J]. Chin J Vector Biol Control, 2022, 33(3): 418-425. DOI:10.11853/j.issn.1003.8280.2022.03.020 |
| [9] |
张芳, 李昊, 田洁, 等. 内蒙古高原长爪沙鼠鼠疫自然疫源地现状分析[J]. 中国地方病防治, 2020, 35(3): 220-222. Zhang F, Li H, Tian J, et al. Analysis on the current situation of Meriones unguiculatus plague natural foci in Inner Mongolia Plateau[J]. Chin J Ctrl Endem Dis, 2020, 35(3): 220-222. |
| [10] |
相蓉, 郭宪国. 二齿新蚤的研究现状[J]. 中国媒介生物学及控制杂志, 2020, 31(5): 623-629. Xiang R, Guo XG. Current research on Neopsylla bidentatiformis[J]. Chin J Vector Biol Control, 2020, 31(5): 623-629. DOI:10.11853/j.issn.1003.8280.2020.05.025 |
| [11] |
岳锡宏, 史深, 袁江玲, 等. 基于不同媒介效能模型的簇鬃客蚤鼠疫媒介效能评价研究[J]. 中国媒介生物学及控制杂志, 2021, 32(5): 564-569. Yue XH, Shi S, Yuan JL, et al. Plague transmission efficiency of Xenopsylla skrjabini based on the vector efficiency evaluation models[J]. Chin J Vector Biol Control, 2021, 32(5): 564-569. DOI:10.11853/j.issn.1003.8280.2021.05.011 |
| [12] |
刘俊, 王建军, 杨秀峰. 内蒙古蚤类的鼠疫流行病学意义[J]. 中国媒介生物学及控制杂志, 2011, 22(6): 576-578. Liu J, Wang JJ, Yang XF. Fleas and the epidemiology of plague in Inner Mongolia, China[J]. Chin J Vector Biol Control, 2011, 22(6): 576-578. |
| [13] |
范蒙光, 李建云, 常子丽, 等. 内蒙古达乌尔黄鼠疫源地主要宿主寄生蚤调查结果分析[J]. 医学动物防制, 2015, 31(9): 945-948. Fan MG, Li JY, Chang ZL, et al. Spermophilus dauricus epidemic focus main hosts parasitical fleas investigation results analysis of Inner Mongolia[J]. J Med Pest Control, 2015, 31(9): 945-948. DOI:10.7629/yxdwfz201509001 |
| [14] |
马丽, 马跃, 张磊, 等. 通辽市2011-2020年动物间鼠疫疫情监测结果分析[J]. 医学动物防制, 2022, 38(8): 801-803. Ma L, Ma Y, Zhang L, et al. Analysis on surveillance results of animal plague in Tongliao from 2011 to 2020[J]. J Med Pest Control, 2022, 38(8): 801-803. DOI:10.7629/yxdwfz202208022 |
| [15] |
刘伯熙, 段然, 王浩珲, 等. 内蒙古自治区不同生态系鼠疫自然疫源地动物鼠疫流行现状及流行风险简析[J]. 中华预防医学杂志, 2022, 56(1): 9-14. Liu BX, Duan R, Wang HH, et al. Analysis on prevalence and epidemic risk of animal plague in different ecological plague foci in Inner Mongolia Autonomous Region[J]. Chin J Prev Med, 2022, 56(1): 9-14. DOI:10.3760/cma.j.cn12150-20211101-01007 |
| [16] |
李仲来, 刘天驰, 牛勇. 锡林郭勒草原布氏田鼠体蚤和巢蚤数量动态及蚤类群落演替[J]. 昆虫学报, 2001(3): 327-331. Li ZL, Liu TC, Niu Y. Studies on dynamics of body and burrow fleas of Microtus brandti and succession of their community[J]. Acta Ento Sin, 2001(3): 327-331. DOI:10.16380/j.kcxb.2001.03.012 |
| [17] |
李忠, 白雪峰. "十一·五"期间内蒙古锡林郭勒盟鼠疫疫源地变化分析[J]. 中国地方病防治杂志, 2012, 27(2): 116-118. Li Z, Bai XF. Analysis of the change of plague foci in Xilin Gol League, Inner Mongolia during the "11th Five Year Plan" Period[J]. Chin J Ctrl Endem Dis, 2012, 27(2): 116-118. |
| [18] |
曹荣季, 邵宗体, 樊荣, 等. 云南省水富市小兽及其体表蚤类的组成与分布[J]. 中国热带医学, 2022, 22(6): 495-499, 516. Cao RJ, Shao ZT, Fan R, et al. Composition and distribution of small mammals and their ectoparasite fleas in Shuifu, Yunnan[J]. China Trop Med, 2022, 22(6): 495-499, 516. DOI:10.13604/j.cnki.46-1064/r.2022.06.02 |
| [19] |
蔡文凤, 罗剑龙, 苏丽琼, 等. 云南省剑川县2008-2017年鼠疫疫源地不同生境小型兽类群落多样性特征[J]. 中国热带医学, 2019, 19(5): 440-443. Cai WF, Luo JL, Su LQ, et al. Community diversity characteristics of small mammals in plage natural focus of Jianchuan county, Yunnan, 2008-2017[J]. China Trop Med, 2019, 19(5): 440-443. DOI:10.13604/j.cnki.46-1064/r.2019.05.09 |
| [20] |
王吉毅. 千岛湖浮游植物群落结构时空分布格局与营养盐的相关性[D]. 哈尔滨: 哈尔滨师范大学, 2022. DOI: 10.27064/d.cnki.ghasu.2022.000988. Wang JY. Correlation between spatial and temporal distribution pattern of phytoplankton community structure and nutrients in Qiandao Lake[D]. Harbin: Harbin Normal University, 2022. DOI: 10.27064/d.cnki.ghasu.2022.000988.(in Chinese) |
| [21] |
陈永明, 闫东, 刘冠纯, 等. 河北省鼠疫自然疫源地长爪沙鼠体蚤多样性研究[J]. 中华卫生杀虫药械, 2018, 24(3): 255-257, 260. Chen YM, Yan D, Liu GC, et al. A diversity of body fleas on Meriones unguiculatus in plague natural epidemic foci of Hebei province[J]. Chin J Hyg Insect Equip, 2018, 24(3): 255-257, 260. DOI:10.19821/j.1671-2781.2018.03.011 |
| [22] |
龚正达, 吴厚永, 段兴德, 等. 云南横断山区蚤类物种多样性的地理分布趋势与重要环境因素的关系[J]. 生物多样性, 2001, 9(4): 319-328. Gong ZD, Wu HY, Duan XD, et al. The relationship between the geographical distribution trends of flea species diversity and the important environmental factor in the Hengduan Mountains, Yunnan[J]. Biodiv Sci, 2001, 9(4): 319-328. DOI:10.3321/j.issn:1005-0094.2001.04.001 |
| [23] |
于建元, 霍敏萍, 郭鹏飞. 锡林浩特市鼠疫疫源地现状的研究[J]. 中国地方病防治杂志, 2012, 27(4): 267-270. Yu JY, Huo MP, Guo PF. Current situation of plague foci in Xilinhot[J]. Chin J Ctrl Endem Dis, 2012, 27(4): 267-270. |
| [24] |
Shi YL, Chi QS, Liu W, et al. Environmental metabolomics reveal geographic variation in aerobic metabolism and metabolic substrates in Mongolian gerbils (Meriones unguiculatus)[J]. Comp Biochem Physiol Part D: Genomics Proteomics, 2015, 14: 42-52. DOI:10.1016/j.cbd.2015.03.001 |