 2022, Vol. 33
2022, Vol. 33扩展功能
文章信息
- 赵美杰, 郭宁宁, 刘起勇, 金华锋, 孟凤霞, 刘小波, 郭玉红
- ZHAO Mei-jie, GUO Ning-ning, LIU Qi-yong, JIN Hua-feng, MENG Feng-xia, LIU Xiao-bo, GUO Yu-hong
- 低温条件下埃及伊蚊生命表研究
- Life table of Aedes aegypti at low temperature
- 中国媒介生物学及控制杂志, 2022, 33(6): 771-775
- Chin J Vector Biol & Control, 2022, 33(6): 771-775
- 10.11853/j.issn.1003.8280.2022.06.001
-
文章历史
- 收稿日期: 2022-07-19




2 东北农业大学生命科学学院, 黑龙江 哈尔滨 150000;
3 山东大学齐鲁医学院公共卫生学院, 山东 济南 250012;
4 东北财经大学经济学院, 辽宁 大连 116025
2 College of Life Science, Northeast Agricultural University, Harbin, Heilongjiang 150000, China;
3 School of Public Health, Cheeloo College of Medicine, Shandong University, Ji'nan, Shandong 250012, China;
4 School of Economics, Dongbei University of Finance and Economics, Dalian, Liaoning 116025, China
埃及伊蚊(Aedes aegypti)是一种家栖型蚊种,是登革热的主要传播媒介,在我国主要分布于云南省边境地区、广东省雷州半岛、海南省火山岩地区、台湾省嘉义县以南及澎湖列岛等地区[1]。生命表(life table)是描述种群死亡及存活情况的一种工具,分为特定年龄生命表和特定时间生命表2种。通过生命表研究,可以了解蚊虫种群动态变化[2-3]。
埃及伊蚊主要生活在热带和亚热带,较多学者对模拟自然条件及高温条件下生命表展开研究。1993年,Focks等[4]提出了容器栖息蚊虫模拟(container-inhabiting mosquito simulation,CIMSiM)生命表模型,旨在阐明饮食对埃及伊蚊存活率、发育时间和繁殖力的影响。2008年Beserra和Castro[5]对巴西帕拉伊巴州埃及伊蚊种群动态研究发现,不同地理种群在相同温度下的生长发育状态和种群生长潜力不同。2013年,Sowilem等[6]研究发现沙特阿拉伯地区的埃及伊蚊种群生长潜力较低。Brady等[7]利用广义相加模型推演埃及伊蚊全球分布图时,发现极端气候下埃及伊蚊生命表数据相当匮乏。近年来,已有学者对18~36 ℃温度下埃及伊蚊种群动态展开研究,结果显示该温度区间内埃及伊蚊能获得较高的长成率[8]。作者早期对10~17 ℃低温下的埃及伊蚊发育做了前期探索,结果表明低于14 ℃时其不能完成世代发育,15~16 ℃时长成率极低,不能完成繁殖。本研究对17 ℃低温下埃及伊蚊生命表展开研究,了解该温度下的种群动态,为埃及伊蚊种群扩散分布研究提供参考依据,进而为相关疾病防控提供基础。
1 材料与方法 1.1 试虫来源及饲养条件 1.1.1 试虫来源海南省儋州市埃及伊蚊地理种群,多年前野外采集并驯化为实验室饲养,经多代繁殖保种于中国疾病预防控制中心传染病预防控制所。
1.1.2 饲养条件生化培养箱设置温度17 ℃(26 ℃对照),相对湿度75%,光照强度8 000 lx,光周期(L∶D)12 h∶12 h。
幼虫于烧杯内盛脱氯水饲养(自来水放置过夜)。每天2次(09:00和21:00)投食(干馒头粉∶猪肝粉=1∶1),投喂量视幼虫大小与数量而定。成蚊饲喂10%蔗糖溶液,羽化3~4 d后饲喂小鼠血,同时提供铺有湿润漏斗状滤纸的烧杯供其产卵。
1.2 实验方法 1.2.1 年龄特征存活率于体视显微镜下挑选24 h内产的饱满卵粒,置于潮湿环境中孵育24 h,然后转入盛有150 ml脱氯水的200 ml烧杯中孵化,每杯200粒卵。每天2次(09:00和21:00)观察记录孵化卵数量,将孵化的幼虫用吸管转移至另一烧杯中继续饲养,观察幼虫生长状况。当第1批孵化幼虫发育至3龄时,判定为本批次卵孵化完毕,停止观察。逐日记录幼虫发育、化蛹和羽化情况。实验设3个重复。计算卵孵化率、幼虫化蛹率、羽化率及成蚊性比(雌∶雄)。
1.2.2 成虫前发育时间初孵幼虫50只一组,每只单独饲养。每天2次(09:00和21:00)投食,并在数字视频显微镜下观察、记录蜕皮幼虫数及蜕皮进入下龄期的时间。实验设3个重复。
1.2.3 成蚊寿命及产卵情况将24 h内化蛹的蚊蛹冲洗后计数100个放入盛水烧杯内待其羽化。3~4 d后,待其全部羽化即开始每3 d饲喂1次小鼠血(持续喂饲24 h),每天2次(09:00和21:00)饲喂10%的蔗糖溶液,并及时清理笼内死亡蚊虫,记录死亡蚊虫性别及数量。每天更换1次滤纸并记录产卵数,直到笼内最后1只蚊虫死亡为止。实验设10个重复。
1.3 数据处理及分析用Excel 2010软件整理所获数据,SPSS 25.0软件进行统计学分析,用GraphPad Prism 8.0.2软件进行绘图。
生命表各项参数计算公式如下(蚊日龄的统计数据从雌、雄蚊开始交配后计算,因此比实际日龄晚3~4 d[9]):

|

|

|

|
(1)各日龄存活率(lx):

|
式中,yx代表日龄为x时的蚊虫存活数;y0代表实验开始时的蚊数;lx代表各日龄组开始时成蚊的存活率。
(2)预期寿命(ex):
|
|
|
式中,Tx代表超过x的各日龄组蚊虫存活率总和;w代表实验种群最后一个日龄组的序数。
(3)净增殖率(R0):
|

|
式中,mx为该每只x日龄雌蚊所产生的雌性后代平均数;Ex为每个x日龄雌蚊所产卵的平均数;s为性别比(雌∶雄);a指卵发育到成蚊过程中存活的成蚊部分;a和s在本文计算中分别采用Walter和Hackes[10]的数值:1.0和0.5;u为开始产卵的日龄组序数。
(4)内禀增长能力(rm):以林昌善[11]改进的公式计算。
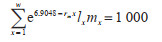
|
(5)世代周期长(T):
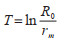
|
(6)增殖有限速率(λ):

|
(7)稳定年龄组配(Px):
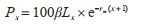
|
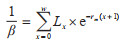
|
式中,Px代表日龄为x~(x+1)的个体在种群中所占百分比;β代表种群在x~(x+1)区间日龄组的相对出生率。
(8)瞬时出生率(B):
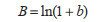
|
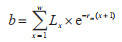
|
式中,b为年(日)龄组为x的个体所占比例。
(9)瞬时死亡率(D):

|
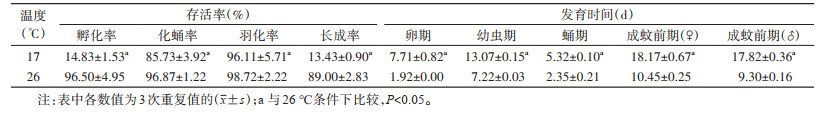
17 ℃时,埃及伊蚊卵的平均孵化率为(14.83±1.53)%,平均化蛹率为(85.73±3.92)%,平均羽化率为(96.11±5.71)%,平均长成率为(13.43±0.90)%。雌、雄成蚊前发育时间分别为(18.17±0.67)和(17.82±0.36)d。从幼虫不同龄期发育时间来看,1、4龄幼虫发育时间较2、3龄幼虫长,其中4龄幼虫发育时间最长,为(5.43±0.93)d。与26 ℃条件下埃及伊蚊各期存活率和发育时间相比,差异均有统计学意义(均P < 0.05)。见表 1。
|
17 ℃时雌性埃及伊蚊平均预期寿命为(21.22±13.73)d,最长可存活98 d,雄性平均预期寿命为(8.00±3.51)d,最长可存活48 d。将成蚊逐日存活状况进行生存分析(图 1),在开始的5 d内,lx相差不大,随着日龄的增长二者差距逐渐拉大,雄性存活率曲线短时间内急剧下降。雌、雄性分别在52和16 d时达到50.00%存活率;98和48 d后完全死亡。

|
| 图 1 17℃下埃及伊蚊成蚊生存分析 Figure 1 Survival analysis of adult Aedes aegypti at 17℃ |
| |
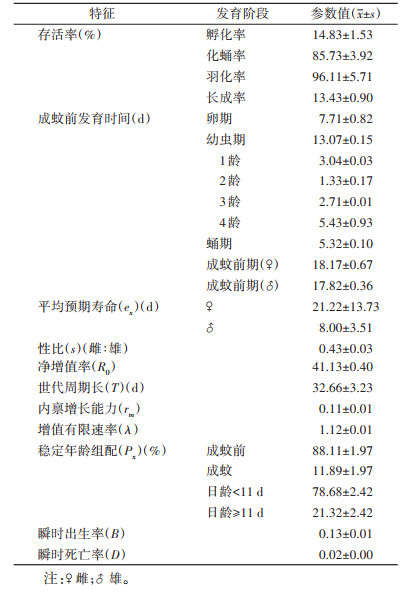
实验中埃及伊蚊在羽化后11 d时首次产卵,末次产卵时间为羽化后第97天,产卵高峰出现在第31~37天。R0为41.13,rm为0.11,T为32.66 d,λ为1.12/d。Px成蚊前期占比为88.11%,成蚊期占比为11.89%,整个成蚊期各日龄组的稳定年龄组配寿命在11 d以内的蚊虫占比为78.68%,寿命在11 d及以上的蚊虫占比为21.32%。B为0.13,D为0.02。17 ℃时埃及伊蚊实验室品系的生命表特征汇总见表 2。
埃及伊蚊分布于热带和亚热带地区。低温条件下的种群动态特征,对预测其分布和扩散具有重要意义。作者前期研究了10~17 ℃条件下埃及伊蚊种群的发育,在≤16 ℃条件下,埃及伊蚊不能完成完整的世代发育。因此,本文对其17 ℃条件下生命表进行研究,以期获得低温下种群动态特征,为预测分布和扩散提供基础。
3.1 低温下埃及伊蚊卵孵化率低,卵期是决定其扩散分布的限制因素Bayoh和Lindsay[12]指出水体温度直接影响埃及伊蚊的生长发育。Lu等[13]研究发现,蚊虫生活的最低温度影响其越冬。本研究结果显示,17 ℃时埃及伊蚊孵化率为(14.83±1.53)%,低于26 ℃时的(96.50±4.95)%;从1龄幼虫开始到成蚊,很少或几乎无死亡,判定卵期是决定其发育的关键阶段。埃及伊蚊分布正逐步扩大,其适生区首先取决于卵期是否能渡过逆境条件,因而可以通过观察卵期在冬季的发育情况,推断其潜在分布。
3.2 低温下埃及伊蚊种群生长潜力较低R0为每一世代蚊虫的种群增值倍数,本研究埃及伊蚊R0为41.13±0.40,表明每一世代的1只雌蚊一生可产下41.13只雌蚊,低于Sowilem等[6]研究的(101.04±38.04)只[温度(25±2)℃;相对湿度(70±10)%;光周期为8 h∶16 h];rm指在既定的物理和生态环境下,具有一定年龄组配的蚊虫种群的瞬时最大增长速率[14],本研究rm为0.11±0.01,表明该世代的每只雌蚊平均每天可产下0.11只雌蚊,低于Beserra和Castro[5]研究的巴西帕拉伊巴州4个地区埃及伊蚊种群的rm(Brejo dos Santos为0.92、Boqueirão为0.78、Itaporanga为0.89、Remígio为0.88)[温度(26±2)℃;光周期为12 h∶12 h]。λ为1.12,指种群数量经过1 d时间所增加的倍数为1.12。各项参数值,说明17 ℃时埃及伊蚊种群数量增长缓慢、繁殖能力较低。
3.3 低温下埃及伊蚊各生活期发育时间较长Tun-Lin等[15]发现低温时埃及伊蚊世代发育时间延长。Kamimura等[16]研究发现,埃及伊蚊及白纹伊蚊的发育周期随温度降低逐渐延长。刘凤梅等[17]研究发现,低温、短光照条件下,中华按蚊(Anopheles sinensis)和淡色库蚊(Culex pipiens pallens)生命周期均有延长。ex能够显示该日龄开始时蚊虫仍能存活的平均天数。本研究发现,雌蚊ex为(21.22±13.73)d,明显高于雄性的(8.00±3.51)d;相比于26 ℃条件下(雌蚊17.14 d,雄蚊9.59 d)[6],雌蚊寿命显著延长,推测为低温下蚊虫代谢活性下降,活动能力下降,各种能量损耗降低所致。Px是指某一年龄段的个体在整个种群生命过程中所占的比例,本研究中寿命在11 d以内的蚊虫占比(78.68±2.42)%,寿命≥11 d的蚊虫占比(21.32±2.42)%。Salazar等[18]在研究埃及伊蚊对登革病毒的传播中发现,当其成蚊寿命延长时,会促进病毒有效传播。Chowell等[19]研究显示登革病毒在蚊虫(宿主)内的潜伏期至少为11 d,该数据说明寿命≥11 d的埃及伊蚊极有可能成为登革病毒的有效传播媒介。当然,埃及伊蚊传播病毒的能力与其寿命、活动能力和叮咬频率均有关系,需要综合考虑各因素的影响。
本研究模拟低温条件对埃及伊蚊种群动态进行研究,填补了极端低温条件下埃及伊蚊生命表特征的研究空白。但本研究是在实验室设定的恒定条件下模拟得出的我国海南地区埃及伊蚊种群的生命表动态,与自然条件下的温度、湿度、光照等存在差异,其生命表各参数尚需进一步研究。
利益冲突 无
| [1] |
刘起勇. 我国登革热流行新趋势、防控挑战及策略分析[J]. 中国媒介生物学及控制杂志, 2020, 31(1): 1-6. Liu QY. Dengue fever in China: New epidemical trend, challenges and strategies for prevention and control[J]. Chin J Vector Biol Control, 2020, 31(1): 1-6. DOI:10.11853/j.issn.1003.8280.2020.01.001 |
| [2] |
Jin YN, Yao JJ, Wang SY, et al. The effect of adding neoadjuvant chemotherapy to concurrent chemoradiotherapy in patients with locoregionally advanced nasopharyngeal carcinoma and undetectable pretreatment Epstein-Barr virus DNA[J]. Transl Oncol, 2017, 10(4): 527-534. DOI:10.1016/j.tranon.2017.03.007 |
| [3] |
Smith C, Khanna R. The development of prophylactic and therapeutic EBV vaccines[M]//Münz C. Epstein Barr Virus Volume 2. Cham: Springer, 2015: 455-473. DOI: 10.1007/978-3-319-22834-1_16.
|
| [4] |
Focks DA, Haile DG, Daniels E, et al. Dynamic life table model for Aedes aegypti (Diptera: Culicidae): Simulation results and validation[J]. J Med Entomol, 1993, 30(6): 1018-1028. DOI:10.1093/jmedent/30.6.1018 |
| [5] |
Beserra EB, Castro FP Jr. Compared biology of populations of Aedes (Stegomyia) aegypti (L.) (Diptera: Culicidae) of Paraíba state, Brazil[J]. Neotrop Entomol, 2008, 37(1): 81-85. DOI:10.1590/s1519-566x2008000100012 |
| [6] |
Sowilem MM, Kamal HA, Khater EI. Life table characteristics of Aedes aegypti (Diptera: Culicidae) from Saudi Arabia[J]. Trop Biomed, 2013, 30(2): 301-314. |
| [7] |
Brady OJ, Johansson MA, Guerra CA, et al. Modelling adult Aedes aegypti and Ae. albopictus survival at different temperatures in laboratory and field settings[J]. Parasit Vectors, 2013, 6: 351. DOI:10.1186/1756-3305-6-351 |
| [8] |
Rueda LM, Patel KJ, Axtell RC, et al. Temperature-dependent development and survival rates of Culex quinquefasciatus and Aedes aegypti (Diptera: Culicidae)[J]. J Med Entomol, 1990, 27(5): 892-898. DOI:10.1093/jmedent/27.5.892 |
| [9] |
王菊生, 王宝鳞. 巴拉巴按蚊实验室种群的动力学一、寿命表[J]. 贵阳医学院学报, 1981, 1-2: 112-119. Wang JS, Wang BL. Kinetics of the laboratory population of Anopheles balabacensis mosquitoes. Ⅰ. Life table[J]. J Guiyang Med Coll, 1981, 1-2: 112-119. DOI:10.19367/j.cnki.1000-2707.1981.z1.023 |
| [10] |
Walter NM, Hacker CS. Variation in life table characteristics among three geographic strains of Culex pipiens quinquefasciatus[J]. J Med Entomol, 1974, 11(5): 541-550. DOI:10.1093/jmedent/11.5.541 |
| [11] |
林昌善. 动物种群数量变动的理论与试验研究Ⅱ. 杂拟谷盗Tribolium confusum (H.) 的内禀增长能力(rm)的研究[J]. 动物学报, 1964, 16(3): 323-338. Lin CS. The theoretical and laboratory studies of animal population dynamics II. A study of innate capacity for increase (rm) of Tribolium confusum (H.) under certain experimental conditions[J]. Acta Zool Sin, 1964, 16(3): 323-338. |
| [12] |
Bayoh MN, Lindsay SW. Effect of temperature on the development of the aquatic stages of Anopheles gambiae sensu stricto (Diptera: Culicidae)[J]. Bull Entomol Res, 2003, 93(5): 375-381. DOI:10.1079/BER2003259 |
| [13] |
Lu L, Lin HL, Tian LW, et al. Time series analysis of dengue fever and weather in Guangzhou, China[J]. BMC Public Health, 2009, 9: 395. DOI:10.1186/1471-2458-9-395 |
| [14] |
胡好远. 种群内禀增长率精确值的简便求法[J]. 安徽农学通报, 2010, 16(3): 173-174. Hu HY. A easy calculation for exact value of the innate rate of increase[J]. Anhui Agric Sci Bull, 2010, 16(3): 173-174. DOI:10.3969/j.issn.1007-7731.2010.03.089 |
| [15] |
Tun-Lin W, Burkot TR, Kay BH. Effects of temperature and larval diet on development rates and survival of the dengue vector Aedes aegypti in north Queensland, Australia[J]. Med Vet Entomol, 2010, 14(1): 31-37. DOI:10.1046/j.1365-2915.2000.00207.x |
| [16] |
Kamimura K, Matsuse IT, Takahashi H, et al. Effect of temperature on the development of Aedes aegypti and Ae. albopictus[J]. Med Entomol Zool, 2002, 53(1): 53-58. DOI:10.7601/mez.53.53_1 |
| [17] |
刘凤梅, 甄天民, 王怀位, 等. 光周期和温度对3种蚊虫寿命影响的观察[J]. 中国寄生虫病防治杂志, 1998, 11(1): 58-61. Liu FM, Zhen TM, Wang HW, et al. Observation on the effect of temperature and photoperiod on the life span of three mosquito species[J]. Chin J Parasit Dis Control, 1998, 11(1): 58-61. |
| [18] |
Salazar MI, Richardson JH, Sánchez-Vargas I, et al. Dengue virus type 2: Replication and tropisms in orally infected Aedes aegypti mosquitoes[J]. BMC Microbiol, 2007, 7: 9. DOI:10.1186/1471-2180-7-9 |
| [19] |
Chowell G, Diaz-Dueñas P, Miller JC, et al. Estimation of the reproduction number of dengue fever from spatial epidemic data[J]. Math Biosci, 2007, 208(2): 571-589. DOI:10.1016/j.mbs.2006.11.011 |