 2022, Vol. 33
2022, Vol. 33扩展功能
文章信息
- 杨怡铃, 何于雯, 李楠, 孟锦昕, 殷建忠, 王静林
- YANG Yi-ling, HE Yu-wen, LI Nan, MENG Jin-xin, YIN Jian-zhong, WANG Jing-lin
- 云南省德宏州东方库蠓中西藏环状病毒的分离鉴定
- Isolation and identification of Tibet orbivirus from Culicoides orientalis collected in Dehong prefecture, Yunnan province, China
- 中国媒介生物学及控制杂志, 2022, 33(4): 580-585
- Chin J Vector Biol & Control, 2022, 33(4): 580-585
- 10.11853/j.issn.1003.8280.2022.04.025
-
文章历史
- 收稿日期: 2022-02-26




2 昆明医科大学公共卫生学院, 云南 昆明 650500
2 School of Public Health, Kunming Medical University, Kunming, Yunnan 650500, China
西藏环状病毒(Tibet orbivirus,TIBOV)是呼肠孤病毒科(Reoviridae)环状病毒属(Orbivirus)中的一名成员[1]。环状病毒属包含22种明确分类的病毒和10种未明确分类的病毒,主要由蠓、蚊、蜱等媒介生物传播致病[2]。其中,环状病毒属的蓝舌病毒(Bluetongue virus,BTV)、非洲马瘟病毒(African horse sickness virus,AHSV)、鹿流行性出血热病毒(Epizootic hemorrhagic disease virus,EHDV)均通过库蠓传播,可感染牛、羊、马和鹿等多种家养与野生反刍动物,动物感染后会出现发热、出血以及呼吸道症状[3-5]。2013年在云南省德宏傣族景颇族自治州(德宏州)芒市芒丙村捕获的蠓中分离出1株TIBOV,并在当地牛和羊的血清中检测到该病毒的中和抗体,说明TIBOV对动物有潜在的致病性,因此应当加强对该病毒的监测与研究[6]。为了证实TIBOV在该地区是否持续存在,本研究于2016年再次在该地区采集蠓样本进行虫媒病毒分离,为当地TIBOV导致动物疾病的研究和防控提供科学依据。
1 材料与方法 1.1 蠓标本采集2016年8月在云南省德宏州芒市芒丙村采用诱蚊灯(功夫小帅,12 V,300 mA,湖北省武汉市生产)在当地的猪圈和羊圈采集标本(18:00至次日07:00)。捕获的蠓在显微镜下进行初步分类后放置-20 ℃条件下冷冻20 min,分装编号后储存在冻存管中,每管大约100只,迅速转移至液氮罐中保存。
1.2 病毒分离取出液氮中冻存的蠓,加入1 ml研磨液(MEM,青、链霉素100 U/ml),放入预冷的研磨器,以25 r/s的频率震荡研磨15 min,4 ℃,6 000×g(8 000 r/min),离心10 min。取200 μl标本研磨的上清液分别接种至长满单层的金黄地鼠肾细胞(BHK-21)和白纹伊蚊卵细胞(C6/36),分别置于含5%CO2的37 ℃及28 ℃培养箱中培养。接种后1~7 d每天观察细胞病变(cytopathic effect,CPE)。2种细胞均盲传3代,3代后无病变则舍弃。
1.3 病毒RNA的提取以及cDNA的制备根据天隆科技病毒RNA提取试剂盒(天隆科技有限公司,西安)的说明,取200 μl病毒上清液,最终洗脱出50 μl核酸。采用Reverse Transcriptase M-MLV(RNase H-)试剂盒(TaKaRa公司,大连)进行RNA反转录合成cDNA。
1.4 病毒的分子生物学鉴定黄病毒属、甲病毒属等特异性引物来源于文献[7-8],参考美国国立生物技术信息中心(NCBI)中5株TIBOV(KU754032、NC027805、KX455493、KF746193、MT793642)S7基因序列设计TIBOV S7节段引物120S7-1F/120S7-1156R(正向引物:5'-GTAAAAATTTGGTGAAGATGGAC-3',反向引物:5'-GTAAGTGTAATTTGGGAAAACGTAT-3');S10节段引物来源于文献[9-10],进行PCR扩增。以cDNA为模板,扩增反应体系为25 μl。PCR反应条件为:94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火1 min,72 ℃延伸30 s,共35个循环;72 ℃中止延伸10 min。1%琼脂糖凝胶电泳用于检测PCR产物条带,阳性产物送至北京擎科生物科技有限公司昆明分公司进行测序。
1.5 蠓基因组DNA的提取与鉴定提取DHC217-11批库蠓基因组DNA:取100 μl研磨液上清,根据天根微量样品基因组DNA试剂盒(天根生化科技有限公司,北京)的说明,最终获得35 μl库蠓基因组DNA。采用细胞色素C氧化酶亚基Ⅰ(COⅠ)基因特异引物(C1-J-1718/C1-N-2191)鉴定库蠓种类[11]。
1.6 基因序列分析从GenBank中下载6株TIBOV以及6株环状病毒属其他病毒的S7和S10基因序列。使用DNAStar(Version:7.1.0)中SeqMan软件对测序结果进行序列拼接;使用DNAStar软件中MegAlign进行病毒基因序列的比对和同源性分析;利用MEGA Ⅹ软件完成基于neighbour-joining(NJ)方法的系统进化树绘制,Bootstrap值为1 000。
2 结果 2.1 DHC217-11病毒株分离2016年8月在云南省德宏州芒市采获蠓约4 400只,分44批进行病毒分离,获得多株阳性分离物,其中1株分离物(编号:DHC217-11)接种BHK-21细胞48 h后产生明显细胞病变(CPE),表现为细胞圆缩、碎裂、脱落;接种C6/36细胞72 h后出现明显CPE,表现为细胞聚体、圆缩、脱落。见图 1。

|
| 注:A BHK-21细胞对照(48 h);B接种DHC217-11的BHK-21细胞(48 h);C C6/36细胞对照(72 h);D接种DHC217-11的C6/36细胞(72 h);→表示BHK-21细胞出现圆缩、碎裂、脱落;↓表示C6/36细胞出现聚体、圆缩、脱落。 图 1 DHC217-11分离病毒株接种BHK-21、C6/36细胞后引起的细胞病变(×200) Figure 1 Cytopathic effect in BHK-21 and C6/36 cells caused by the infection with DHC217-11 isolate (×200) |
| |
采用TIBOV S7及S10节段引物对DHC217-11进行反转录聚合酶链式反应(reverse transcription-polymerase chain reaction,RT-PCR),分别在1 085和723 bp出现明显的电泳条带,而黄病毒属和甲病毒属引物扩增均为阴性。测序后经BLAST比对发现,新毒株DHC217-11 S7基因序列与TIBOV DH13C120核苷酸序列具有99.5%同源性;S10基因序列与TIBOV SX-2017a核苷酸具有97.1%同源性,初步鉴定库蠓中新分离病毒(DHC217-11)为TIBOV。
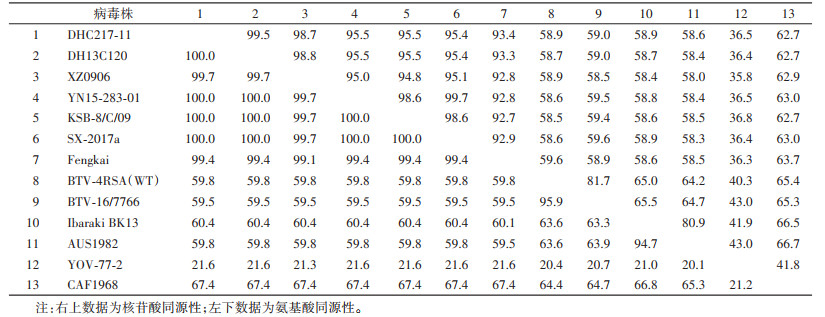
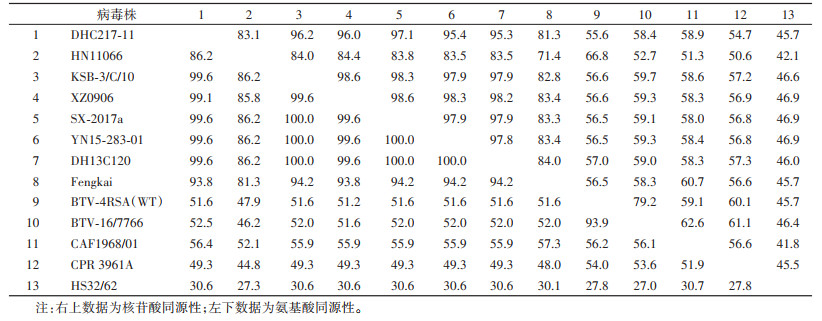
2.3 新分离病毒株S7和S10基因序列核苷酸和氨基酸同源性分析从GenBank下载来自不同国家、地区、媒介宿主分离的环状病毒属代表性成员的S7和S10基因序列,与DHC217-11进行氨基酸与核苷酸的同源性分析(表 1、2),结果显示库蠓新分离DHC217-11株S7基因序列与2013年德宏州芒市采集蠓中分离的DH13C120同源性最高,核苷酸和氨基酸同源性分别为99.5%和100%;其次与中国西藏自治区(西藏)、广东省、云南省西双版纳傣族自治州(西双版纳州)及日本等地分离的TIBOV核苷酸和氨基酸同源性分别在93.4%~98.7%和99.4%~100%;与澳大利亚、日本等地分离的EHDV,南非分离的BTV,云南省分离的云南环状病毒(Yunnan orbivirus,YOUV)核苷酸和氨基酸同源性较低,分别低于62.7%和67.4%。DHC217-11株S10基因序列与2017年从云南省西双版纳州采集蚊虫分离的SX-2017a同源性最高,核苷酸和氨基酸同源性分别为97.1%和99.6%;与中国西藏、广东省、云南省西双版纳州及日本等地分离的TIBOV核苷酸和氨基酸同源性在81.3%~96.2%和86.2%~99.6%;与澳大利亚分离的EHDV,南非分离的BTV、AHSV,中国云南省分离的YOUV等的核苷酸和氨基酸同源性较低,均低于58.9%和56.4%。
|
|
将库蠓新分离病毒DHC217-11的S7和S10基因序列与GenBank中下载的6株TIBOV,2株BTV、2株EHDV、1株YOUV、1株Pata病毒的相应基因序列一起采用MEGA X软件构建系统进化树(图 2A、B),结果显示,在S7和S10进化树上,DHC217-11与TIBOV属于同一进化分支,而与BTV-4RSA(WT)、BTV-16/7766、EHDV、YOV-77-2和AUS1982等环状病毒属的其他成员位于不同进化分支。

|
| 注:●表示本研究分离病毒株;A表示DHC217-11 S7基因(1 085 bp)系统进化分析;B表示DHC217-11 S10基因(723 bp)系统进化分析。 图 2 DHC217-11分离株S7和S10基因系统进化分析 Figure 2 Phylogenetic analysis of S7 and S10 gene in the DHC217-11 isolate |
| |
在解剖镜下观察DHC217-11批库蠓,其翅形态与东方库蠓相似(图 3A)。采用COⅠ特异引物扩增得到DHC217-11批库蠓COⅠ基因序列,与GenBank中来自不同地区、不同年份的蠓COⅠ基因构建系统进化树(图 3B),结果显示,DHC217-11批库蠓与东方库蠓位于同一进化分支,其同源性为99.8%。

|
| 注:●本研究中DHC 217-11批库蠓;A DHC217-11批库蠓翅面形态;B DHC217-11批库蠓细胞色素C氧化酶亚基Ⅰ基因系统进化分析。 图 3 DHC217-11批库蠓形态及其分子生物学鉴定 Figure 3 Morphology and molecular biological identification of Culicoides from batch DHC217-11 |
| |
我国学者自2009年首次在西藏捕获的圆斑按蚊(Anopheles maculatus)中分离出TIBOV(XZ0906株)以来[12],研究者曾陆续在湖南省长沙市捕获的三带喙库蚊(Culex tritaeniorhynchus)、广东省的致倦库蚊(Cx. pipiens quinquefasciatus)、海南省的野生蠓、云南省德宏州和师宗县的库蠓中分离出新的TIBOV[6, 13-16]。迄今为止,TIBOV在我国北方地区被分离出的文献还未见报道。可见TIBOV广泛分布在我国西南、中东部以及南部沿海地区。本研究再次从我国云南省德宏州芒市采获的东方库蠓分离出1株病毒,能引起BHK-21细胞(48 h)、C6/36细胞(72 h)产生明显的CPE,其CPE特点与2013年云南省德宏州芒市采获蠓分离的DH13C120病毒株相似[6]。采用TIBOV S7和S10基因特异性引物扩增均为阳性,提示东方库蠓新分离病毒DHC217-11可能为环状病毒属的TIBOV。
序列分析结果显示,DHC217-11株病毒S7和S10基因序列与中国西藏、广东省以及日本等地分离的TIBOV位于同一进化分支内,它们之间的核苷酸与氨基酸同源性也较高;与环状病毒属的其他代表性成员位于不同进化分支,它们之间的核苷酸和氨基酸同源性较低,提示2016年云南省德宏州芒市采集东方库蠓分离的病毒DHC217-11为TIBOV。进一步分析发现,新分离病毒株DHC217-11与2013年云南省德宏州芒市采获蠓分离的DH13C120株病毒的S7基因序列核苷酸和氨基酸同源性分别为99.5%和100%,S10基因序列核苷酸和氨基酸同源性分别为95.3%和99.6%,DHC217-11与DH13C120遗传关系最为密切,表明DH13C120病毒株在当地的自然界持续存在。
库蠓是环状病毒属病毒的重要传播媒介,可传播多种疾病,如蓝舌病、鹿流行性出血热、非洲马瘟病、奥西罗热、水泡口炎等,不仅危害人类和动物的健康,而且给畜牧产业带来严重的经济损失[17]。既往研究者在云南省师宗县采获的库蠓和牛血清中分离和检测到TIBOV,表明该病毒可以感染当地的家养动物[18]。本研究在云南省德宏州芒市牛圈采获的东方库蠓中分离到TIBOV,提示东方库蠓可能作为一种TIBOV潜在的传播媒介。东方库蠓广泛分布于我国福建、台湾、海南、云南、西藏等地区,提示这些地区有发生TIBOV流行的风险,今后应加强该地区TIBOV的调查研究,开展防蠓灭蠓活动,减少该病毒给人兽健康带来的危害[19]。
利益冲突 无
| [1] |
Kapoor A, Tesh RB, Duraisamy R, et al. A novel mosquito-borne Orbivirus species found in South-east Asia[J]. J Gen Virol, 2013, 94(Pt 5): 1051-1057. DOI:10.1099/vir.0.046748-0 |
| [2] |
Roy P. Orbivirus structure and assembly[J]. Virology, 1996, 216(1): 1-11. DOI:10.1006/viro.1996.0028 |
| [3] |
李占鸿, 宋子昂, 廖德芳, 等. 云南省蓝舌病病毒血清7型毒株的分离与基因组序列分析[J]. 畜牧兽医学报, 2021, 52(5): 1337-1348. Li ZH, Song ZA, Liao DF, et al. Whole genome sequence analyses of Bluetongue virus serotype 7 strain isolated from Yunnan province[J]. Acta Vet Zootech Sin, 2021, 52(5): 1337-1348. DOI:10.11843/j.issn.0366-6964.2021.05.019 |
| [4] |
朱建波, 董国栋, 杨振兴, 等. 非洲马瘟在东南亚的暴发及其对云南马属动物的威胁[J]. 云南畜牧兽医, 2021(2): 18-21. Zhu JB, Dong GD, Yang ZX, et al. The outbreak of African horse sickness in Southeast Asia and its threats to equus in Yunnan[J]. Yunnan J Anim Sci Vet, 2021(2): 18-21. |
| [5] |
黄超华, 花群俊, 吴江, 等. 鹿流行性出血热病毒核心样颗粒的制备与鉴定[J]. 中国兽医科学, 2021, 51(8): 1023-1028. Huang CH, Hua QY, Wu J, et al. Identification and synthesis of core-like particles of epizootic hemorrhagic disease virus[J]. Chin Vet Sci, 2021, 51(8): 1023-1028. DOI:10.16656/j.issn.1673-4696.2021.0141 |
| [6] |
Wang JL, Li HC, He YW, et al. Isolation of Tibet orbivirus from Culicoides and associated infections in livestock in Yunnan, China[J]. Virol J, 2017, 14(1): 105. DOI:10.1186/s12985-017-0774-9 |
| [7] |
Kuno G. Universal diagnostic RT-PCR protocol for arboviruses[J]. J Virol Methods, 1998, 72(1): 27-41. DOI:10.1016/s0166-0934(98)00003-2 |
| [8] |
Pfeffer M, Proebster B, Kinney RM, et al. Genus-specific detection of alphaviruses by a semi-nested reverse transcription-polymerase chain reaction[J]. Am J Trop Med Hyg, 1997, 57(6): 709-718. DOI:10.4269/ajtmh.1997.57.709 |
| [9] |
李楠, 何于雯, 孟锦昕, 等. 西藏环状病毒荧光定量RT-PCR检测方法的建立[J]. 中国兽医科学, 2020, 50(11): 1360-1364. Li N, He YW, Meng JX, et al. Development of the detection method of real-time RT-PCR assay for Tibet orbivirus[J]. Chin Vet Sci, 2020, 50(11): 1360-1364. DOI:10.16656/j.issn.1673-4696.2020.0174 |
| [10] |
元正菊, 李楠, 何于雯, 等. 西藏环状病毒RT-PCR检测方法的建立[J]. 云南畜牧兽医, 2020(3): 10-13. Yuan ZJ, Li N, He YW, et al. Development of the detection method of RT-PCR assay for Tibet orbivirus[J]. Yunnan J Anim Sci Vet Med, 2020(3): 10-13. DOI:10.3969/j.issn.1005-1341.2020.03.004 |
| [11] |
Dallas JF, Cruickshank RH, Linton YM, et al. Phylogenetic status and matrilineal structure of the biting midge, Culicoides imicola, in Portugal, Rhodes and Israel[J]. Med Vet Entomol, 2003, 17(4): 379-387. DOI:10.1111/j.1365-2915.2003.00454.x |
| [12] |
Li MH, Zheng YY, Zhao GY, et al. Tibet orbivirus, a novel Orbivirus species isolated from Anopheles maculatus mosquitoes in Tibet, China[J]. PLoS One, 2014, 9(2): e88738. DOI:10.1371/journal.pone.0088738 |
| [13] |
范娜, 曹玉玺, 付士红, 等. 三带喙库蚊中分离的西藏环状病毒全基因序列测定及分析[J]. 病毒学报, 2018, 34(5): 625-637. Fan N, Cao YX, Fu SH, et al. Genome sequencing and analyses of the Tibet orbivirus isolated from Culex tritaeniorhynchus[J]. Chin J Virol, 2018, 34(5): 625-637. DOI:10.13242/j.cnki.bingduxuebao.003452 |
| [14] |
Wu D, Tan QQ, Zhang H, et al. Genomic and biological features of a novel orbivirus isolated from mosquitoes, in China[J]. Virus Res, 2020, 285: 197990. DOI:10.1016/j.virusres.2020.197990 |
| [15] |
王琴燕, 付士红, 孙定炜, 等. 海南省野生蠓虫西藏环状病毒的研究[J]. 中国媒介生物学及控制杂志, 2021, 32(4): 415-421. Wang QY, Fu SH, Sun DW, et al. A study of wild midges carrying Tibet orbivirus in Hainan province, China[J]. Chin J Vector Biol Control, 2021, 32(4): 415-421. DOI:10.11853/j.issn.1003.8280.2021.04.006 |
| [16] |
杨振兴, 谢佳芮, 李占鸿, 等. 1株分离自云南省师宗县库蠓的新型西藏环状病毒[J]. 畜牧兽医学报, 2021, 52(1): 166-176. Yang ZX, Xie JR, Li ZH, et al. Isolation and identification of a novel type Tibet orbivirus strain from Culicoides collected in Shizong county of Yunnan province[J]. Acta Vet Zootech Sin, 2021, 52(1): 166-176. DOI:10.11843/j.issn.0366-6964.2021.01.017 |
| [17] |
何于雯, 李钊, 孟锦昕, 等. 云南省江城县五黑阿蠓形态学鉴定及ITS-1和COⅠ基因序列分析[J]. 中国国境卫生检疫杂志, 2020, 43(5): 321-324. He YW, Li Z, Meng JX, et al. Morphological identification and ITS-1 and COⅠ gene sequence analysis of Alluaudomyia quinquepicina in Jiangcheng county, Yunnan province[J]. Chin J Front Health Quart, 2020, 43(5): 321-324. DOI:10.16408/j.1004-9770.2020.05.006 |
| [18] |
李占鸿, 杨振兴, 谢佳芮, 等. 首次在云南省哨兵牛上分离出一种新型西藏环状病毒[J]. 病毒学报, 2021, 37(4): 922-932. Li ZH, Yang ZX, Xie JR, et al. A novel serotype of Tibet orbivirus isolated from sentinel cattle in Yunnan province, China[J]. Chin J Virol, 2021, 37(4): 922-932. DOI:10.13242/j.cnki.bingduxuebao.004010 |
| [19] |
虞以新. 中国蠓科昆虫(昆虫纲, 双翅目)[M]. 北京: 军事医学科学出版社, 2006: 962-963. Yu YX. Ceratopogonidae of China (Insecta, diptera)[M]. Beijing: Military Medical Science Press, 2006: 962-963. |