 2022, Vol. 33
2022, Vol. 33扩展功能
文章信息
- 孙元超, 刘凯, 姚晓燕, 寸得娇, 田娜, 张仪, 王飞, 李兰花
- SUN Yuan-chao, LIU Kai, YAO Xiao-yan, CUN De-jiao, TIAN Na, ZHANG Yi, WANG Fei, LI Lan-hua
- 基于MaxEnt模型对我国纹沼螺潜在分布区的预估研究
- Maximum entropy model-based projection of potential distribution areas of Parafossarulus striatulus in China
- 中国媒介生物学及控制杂志, 2022, 33(4): 555-561
- Chin J Vector Biol & Control, 2022, 33(4): 555-561
- 10.11853/j.issn.1003.8280.2022.04.021
-
文章历史
- 收稿日期: 2022-02-22




2 中国疾病预防控制中心寄生虫病预防控制所媒传热带病室, 上海 200025
2 National Institute of Parasitic Diseases, Chinese Center for Disease Control and Prevention, Shanghai 200025, China
纹沼螺(Parafossarulus striatulus)作为豆螺科的一种,是我国华支睾吸虫(Clonorchis sinensis)重要的第一中间宿主,其生活范围比较广泛,常见的有溪流、湖泊、池塘以及稻田等,华支睾吸虫的虫卵可经过第一中间宿主纹沼螺,第二中间宿主鱼虾以及终宿主人、猫等逐渐发育成成虫,进而对终宿主的肝脏造成损害[1]。掌握纹沼螺在我国的分布情况将对华支睾吸虫病的防控工作有重要意义。因此,本研究应用最大熵模型(MaxEnt)结合纹沼螺已有的分布数据和环境因子信息,分析相关气候因子及生物环境因子对我国纹沼螺分布的影响,并预估气候变化可能对未来纹沼螺潜在适生区分布的影响,研究结果可为华支睾吸虫病防制工作提供参考。
第六次国际耦合模式比较计划(CMIP6)不同于CMIP5中的典型浓度路径(RCPs)情景,CMIP6中的情景是不同共享社会经济路径(SSPs)和RCPs的组合情景,其含义在于包含了未来社会经济发展。SSP-RCP情景主要包括7个模式,分别是SSP1-1.9、SSP1-2.6、SSP4-3.4、SSP2-4.5、SSP4-6.0、SSP3-7.0和SSP5-8.5。本次研究主要运用其中2个情景模式:SSP1-2.6属于低强迫情景,2100年辐射强迫稳定在约2.6 W/m2,该情景考虑了未来全球森林覆盖面积的增加并伴随大量的土地利用变化,通过模型的综合评估,形成了低脆弱性、低减缓挑战的特征,符合SSPs情景;SSP2-4.5属于中等辐射强迫情景,2100年辐射强迫稳定在约4.5 W/m2,由于土地利用和气溶胶路径并不极端,仅代表结合了1个中等社会脆弱性和中等辐射强迫的情景[2]。
1 材料与方法 1.1 纹沼螺分布点的获取以“纹沼螺”为检索词在中国知网和万方数据知识服务平台(万方)数据库中进行全文检索,同时,以“(Parafossarulus striatulus)AND(China)”为检索词在PubMed和Web of Science数据库中进行检索,检索日期为1900年1月1日至2021年6月15日,从包含我国纹沼螺分布的文献中提取纹沼螺地理位置及坐标信息,若文献中只有地理位置,则利用百度地图的“坐标转换”功能将地理位置信息转换为分布点的坐标。同时从世界生物多样性信息交换平台(GBIF)中获取纹沼螺其他分布点信息。
若文献收集的纹沼螺地理分布点过于集中,会造成MaxEnt模型过拟合、不准确,为避免上述问题发生,运用ArcGIS 10.1软件缓冲区筛选功能,半径设置为5 km,筛除缓冲区内的多余分布点,使每个缓冲区中仅保留1个分布点用作预估工作。
1.2 气候因子和地理因子的获取以及分析在全球气候数据网站(https://www.worldclim.org/)中下载当前气候模式下19个气候因子,空间分辨率选择5 min;同时,分别下载SSP126气候情景和SSP245气候情景下,未来2041-2060年和2061-2080年的19个气候因子。其他地理因子为海拔、坡向、坡度、植被覆盖率、有机碳、土壤酸碱度和水体指数,从航天飞机雷达地形测绘任务(shuttle radar topography mission,SRTM)数据(http://srtm.Csi.Cgiar.org/)中可获得所需海拔数据,运用ArcGIS 10.1软件“表面分析”功能分析并处理上述海拔数据可获得所需坡向和坡度数据,植被覆盖率数据经地理空间数据云平台(www.gscloud.cn)查询获得;水体指数可运用遥感影像的特定波段进行差值处理获得。
在maxent 3.4.1软件中导入缓冲区筛选后纹沼螺分布点、19个当前气候因子以及7个地理因子数据,运行1次后由结果获得所有因子贡献率,进而运用ArcGIS 10.1软件“采样”功能提取全部纹沼螺分布点的19个气候因子信息,再运用SPSS 26.0软件中“相关性分析”功能处理分析19个气候因子,从而得到19个气候因子之间的相关系数|r|(|r|越接近于1,表明两变量相关程度越高)。为避免模型因气候因子多重共线性发生过拟合,当出现2个气候因子的r > 0.9时,表明2个气候因子相关程度高,因此选取贡献率较大的气候因子用作模型构建[3],贡献率较大表明对纹沼螺分布影响较大且建立的预估模型更具准确性。
1.3 当前我国纹沼螺的适生区分布预估在maxent 3.4.1软件中准确导入筛选后的当前气候因子、地理因子和分布点信息,用作构建模型的数据训练集设定为75.0%,系统收缩阈值设定为10-5,最大迭代次数设定为500,交叉验证重复运算10次构建模型。将MaxEnt模型运算结果导入ArcGIS 10.1软件中,再运用“自然断点法”将所得适宜度指数划分为4个层次,从而得到当前我国纹沼螺适生区分布情况图。
受试者工作特征曲线(ROC)作为评价MaxEnt模型预估精确度的标准,若ROC曲线下的面积(AUC)在0.8~0.9(不包括0.9),则可认定预估良好[4-6],AUC≥0.9则可认定预估很好。
1.4 不同气候模式下气候变化对未来纹沼螺在我国的分布影响在SSP126情景模式下,结合2041-2060年气候因子和其他地理因子,运用上述建立的MaxEnt模型,预估2041-2060年我国纹沼螺适生区[7];结合2061-2080年气候因子和其他地理因子,运用上述建立的MaxEnt模型,预估2041-2060年我国纹沼螺适生区。SSP245情景模式的模型预估工作与SSP126相同[8]。
应用ArcGIS 10.1软件分析MaxEnt模型所得预估结果,并结合中国矢量地图得到我国纹沼螺潜在适生区分布图,同时计算适生区相关面积。在制作适生区分布图的“重分类”步骤中,运用“自然间断点法”将我国纹沼螺生境适宜指数分为4个等级:非适生区分布概率值< 0.08,低适生区0.08~0.26(不含0.26),中适生区0.26~0.49,高适生区 > 0.49。
2 结果 2.1 纹沼螺分布点最终共获得1 004篇中英文文献,从中收集到225个纹沼螺分布点信息,同时从GBIF中获取9个纹沼螺分布点信息,经设置5 km的缓冲区后,仅保留198个分布点用于模型构建。
2.2 环境因子对纹沼螺分布的影响最终筛选获得11个气候因子和7个地理因子用作构建模型,各环境因子贡献率见表 1,其中海拔(贡献率51.42%)、水体指数(贡献率11.23%)、最湿月降水量(贡献率8.71%)和最干月降水量(贡献率6.82%)对纹沼螺分布贡献率高。
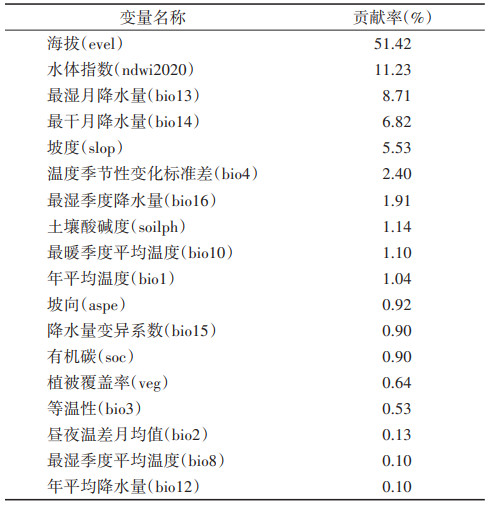
|
对纹沼螺分布概率影响贡献率较高的环境因子响应曲线见图 1。响应曲线表明,随着海拔的升高,纹沼螺分布概率先平缓然后逐渐下降,其中在-838.1~109.9 m时,纹沼螺分布概率高且超过50%;随着水体指数的增加,纹沼螺分布概率先上升后下降再上升最后轻微下降,总体呈上升趋势,其中在-0.527~-0.344和0.321~0.988时,纹沼螺分布概率高且超过50%;随着最湿月降水量的增加,纹沼螺分布概率先上升后下降,其中在183.4~337.2 mm时,纹沼螺分布概率高且超过50%;随着最干月降水量的增加,纹沼螺分布概率先上升后下降,其中在17.2~44.2 mm时,纹沼螺分布概率高且超过50%。

|
| 图 1 纹沼螺分布概率与4种重要环境因子响应曲线 Figure 1 Response curve of probability of Parafossarulus striatulus distribution and four major environmental factors |
| |
MaxEnt模型预估结果显示,AUC为0.938,说明本模型预估精度高。预估结果显示,当前我国纹沼螺高适生区面积为37.37万km2,主要分布在广西、广东、湖南、江西、湖北、安徽、江苏、山东、辽宁等省(自治区);中适生区面积为61.60万km2,主要分布在广西、湖南、湖北、四川、山东、河北等省(自治区);低适生区面积为141.26万km2,主要分布在广西、广东、四川、湖北、河南、河北、辽宁、吉林、黑龙江等省(自治区);非适生区面积为712.43万km2,主要分布在新疆、西藏、青海、甘肃、内蒙古、宁夏、山西等省(自治区)。见图 2。

|
| 图 2 当前我国纹沼螺适生区分布 Figure 2 Present distribution of suitable habitats of Parafossarulus striatulus in China |
| |
MaxEnt模型预估结果显示(表 2),SSP126情景下,2041-2060年我国纹沼螺高适生区面积比当前增加91.00万km2,中适生区面积将增加64.53万km2,低适生区面积将增加101.55万km2,而非适生区面积将减少267.09万km2。2061-2080年我国纹沼螺高适生区面积比当前将增加93.82万km2,中适生区面积将增加67.35万km2,低适生区面积将增加100.80万km2,而非适生区面积将减少261.98万km2。适生区分布情况见图 3。

|

|
| 图 3 SSP126情景模式下不同时间我国纹沼螺适生区分布 Figure 3 Distribution of suitable habitats of Parafossarulus striatulus in China at different times under shared socioeconomic pathway 126 |
| |
MaxEnt模型预估结果显示(表 3),SSP245情景下,2041-2060年我国纹沼螺高适生区面积比当前将增加41.22万km2,中适生区面积将增加76.73万km2,低适生区面积将增加191.62万km2,而非适生区面积将减少309.46万km2。2061-2080年我国纹沼螺高适生区面积比当前将增加49.44万km2,中适生区面积将增加71.98万km2,低适生区面积将增加196.86万km2,而非适生区面积将减少318.26万km2。适生区分布情况见图 4。

|

|
| 图 4 SSP245情景模式下不同时间我国纹沼螺适生区分布 Figure 4 Distribution of suitable habitats of Parafossarulus striatulus in China at different times under shared socioeconomic pathway 245 |
| |
MaxEnt模型是物种分布模型中常用且准确性高的一种模型,可将物种分布点和环境因子结合在一个系统,最大程度地呈现物种的适生区分布,考虑避免该模型拟合过大的问题,可将参数FC和RM分别设置为LQHP和3,此时该模型的拟合程度较低[9],因此本研究选取上述参数进行。鉴于以往学者未曾涉及纹沼螺适生区预估的研究,本研究运用MaxEnt模型分析了环境因子对我国纹沼螺分布的影响,并将不同情景模式下气候变化对其分布的影响进行了预估。分析结果表明,环境因子中,海拔、水体指数、最湿月降水量和最干月降水量对纹沼螺分布的影响较大,未来气候变化使纹沼螺的适生区分布不同程度扩大。
纹沼螺为腹足纲螺类生物,适合的温度范围是其正常的生长、繁殖和取食活动的必要条件[10],在一定温度范围内,螺类的月增重、母螺产仔数量[11]、螺卵孵出时间[12]均与温度呈正相关趋势。预估结果表明,海拔对于纹沼螺分布影响极大,当海拔在-838.1~109.9 m时,纹沼螺适生程度高,考虑海拔可能影响温度,气温一般沿海拔呈递减趋势,生态学代谢理论(MTE)表明,高温可加快生物新陈代谢速率,加快某地区物种形成速率,进而保持较高的物种丰富度[13],结合图 2可以发现中高适生区集中分布在我国东南地区和华中地区,该地区海拔多 < 100 m,且该地区多为亚热带季风气候(最冷月平均温度在0~15 ℃,最热月平均温度 > 22 ℃)和温带季风气候(最冷月平均温度低于0 ℃,最热月平均温度 > 22 ℃)[14],结合图 3、4可以发现,中高适生区的范围扩大到了东北地区,且该地区也属于温带季风气候区域,Dudgeon曾指出,热带亚热带河溪中的水生生物多样性要明显高于温带地区[15],当气温在15~35 ℃时,螺的发育速率随温度逐渐加快,最适宜繁衍分布[16]。因此温度适宜的亚热带季风区和温带季风区纹沼螺分布概率较高,而影响分布概率的最终因素则是海拔,适生区向北扩张也得到了解释。但图 3、4未来适生区表明,新疆维吾尔自治区以及内蒙古自治区出现了新增适生区,结合图 2可以发现,该新增适生区为低适生区新增,可能为白杨河水库区域和呼伦湖区域,水体生物相对较多,且受低温条件影响,人类活动较少[17]。图 3、4未来适生区的结果还表明,新增中高适生区多分布在我国华北和东北地区,原因可能为人类活动引发的全球气候变暖,有研究表明,1906-2005年近100年的温度变化线性上升趋势为0.74 ℃[18],且这种气温上升的趋势仍在继续,同样有研究表明,我国亚热带季风区域和温带季风区域的范围将会扩大[19],因此我国华北和东北地区在温度逐渐升高的条件下,相比较当前而言更适合纹沼螺生存。在华支睾吸虫防控工作中,应重点考虑我国温度变化趋势对纹沼螺分布的影响。
预估结果表明,纹沼螺分布概率也受水体指数、最湿月降水量和最干月降水量的影响,可能为以下原因:纹沼螺作为华支睾吸虫的第一中间宿主,在我国血吸虫病流行区多靠近水源分布,无论幼螺还是成螺都需要在水中或湿润地区生存,因此水域充沛地区更有利于纹沼螺繁衍分布。我国东南地区地处亚热带季风气候区,平均年降水量在800~1 500 mm,华北和东北地区地处温带季风气候,平均年降水量在400~800 mm,由于纹沼螺的分布受地下水量的影响[20],降雨量多的地区地下水量也多,土壤湿度也就越大,纹沼螺适生程度自然就高[21]。根据现有的研究表明,基于政府间气候变化专门委员会(IPCC)的A1B情景,截止到21世纪中期,绝大部分地区降水将呈现增长趋势,夏季变化在-0.1~1.1 mm/d,冬季变化在-0.2~0.2 mm/d,只有西部极少地区呈现减少趋势[22],该降水变化与本研究适生区变化的趋势相符合。
基于2种情景模式的分析可以得出:SSP126低强迫模式下,由于未来全球森林覆盖面积的增加,未来2041-2060年和2061-2080年2个周期内的纹沼螺适生区大范围增加,而在SSP245中等强迫模式下未来2041-2060年和2061-2080年2个周期内的纹沼螺适生区小范围增加,2个情景模式的总体变化趋势相一致,因此结合上述预估的结果,要加强在纹沼螺适生区的知识普及以及健康教育工作,以提高人们对于预防感染华支睾吸虫的意识,从而更好地完成防疫工作。
本次研究仍有一些不足之处,首先,本研究仅从文献中提取纹沼螺的分布点,分布点信息可能不全面,建议未来建立针对螺类的系统监测体系,以更准确地掌握纹沼螺及其他有害螺类的分布范围;其次,本次研究仅分析了气候和地理因子对纹沼螺分布的影响,未考虑人类社会活动及自然灾害的影响。因此,未来研究可纳入新的影响因素进一步用于纹沼螺适生区的分析。
利益冲突 无
| [1] |
江颖. 我国华支睾吸虫中间宿主螺类分子系统发育研究[D]. 北京: 中国疾病预防控制中心, 2016. Jiang Y. Molecular phylogeny on intermediate host snails of Clonorchis sinensis in China[D]. Beijing: Chinese Center for Disease Control and Prevention, 2016. (in Chinese) |
| [2] |
姜彤, 吕嫣冉, 黄金龙. CMIP6模式新情景(SSP-RCP)概述及其在淮河流域的应用[J]. 气象科技进展, 2020, 10(5): 102-109. Jiang T, Lyu YR, Huang JL. New scenarios of CMIP6 model (SSP-RCP) and its application in the Huaihe River basin[J]. Adv Meteor Sci Technol, 2020, 10(5): 102-109. DOI:10.3969/j.issn.2095-1973.2020.05.016 |
| [3] |
叶永昌, 周广胜, 殷晓洁. 1961-2010年内蒙古草原植被分布和生产力变化: 基于MaxEnt模型和综合模型的模拟分析[J]. 生态学报, 2016, 36(15): 4718-4728. Ye YC, Zhou GS, Yin XJ. Changes in distribution and productivity of steppe vegetation in Inner Mongolia during 1961 to 2010:Analysis based on MaxEnt model and synthetic model[J]. Acta Ecol Sin, 2016, 36(15): 4718-4728. DOI:10.5846/stxb201412302599 |
| [4] |
王雨生, 王召海, 邢汉发, 等. 基于MaxEnt模型的珙桐在中国潜在适生区预测[J]. 生态学杂志, 2019, 38(4): 1230-1237. Wang YS, Wang ZH, Xing HF, et al. Prediction of potential suitable distribution of Davidia involucrata baill in China based on MaxEnt[J]. Chin J Ecol, 2019, 38(4): 1230-1237. DOI:10.13292/j.1000-4890.201904.024 |
| [5] |
王茹琳, 王闫利, 陈东东, 等. 基于MaxEnt模型的柑橘木虱在西南地区潜在分布模拟[J]. 植物保护, 2021, 47(1): 84-90, 96. Wang RL, Wang YL, Chen DD, et al. Analysis of the potential distribution of the Asian citrus psyllid, Diaphorina citri Kuwayama in Southwest China using the MaxEnt model[J]. Plant Prot, 2021, 47(1): 84-90, 96. DOI:10.16688/j.zwbh.2019592 |
| [6] |
何文文, 伍军, 温丽翠, 等. MaxEnt模型对新疆盾糙璃眼蜱生境适应性预测[J]. 中国动物检疫, 2022, 39(2): 45-51. He WW, Wu J, Wen LC, et al. Prediction to the habitat adaptability of Hyalomma scupense in Xinjiang by MaxEnt model[J]. China Anim Health Insp, 2022, 39(2): 45-51. DOI:10.3969/j.issn.1005-944X.2022.02.009 |
| [7] |
杨绚, 李栋梁, 汤绪. 基于CMIP5多模式集合资料的中国气温和降水预估及概率分析[J]. 中国沙漠, 2014, 34(3): 795-804. Yang X, Li DL, Tang X. Probability assessment of temperature and precipitation over China by CMIP5 multi-model Ensemble[J]. J Desert Res, 2014, 34(3): 795-804. DOI:10.7522/j.issn.1000-694X.2013.00381 |
| [8] |
张蕾, 黄大鹏, 杨冰韵. RCP4.5情景下中国人口对高温暴露度预估研究[J]. 地理研究, 2016, 35(12): 2238-2248. Zhang L, Huang DP, Yang BY. Future population exposure to high temperature in China under RCP4.5 scenario[J]. Geogr Res, 2016, 35(12): 2238-2248. DOI:10.11821/dlyj201612004 |
| [9] |
贾翔, 王超, 金慧, 等. 基于优化的MaxEnt模型评价红松适宜分布区[J]. 生态学杂志, 2019, 38(8): 2570-2576. Jia X, Wang C, Jin H, et al. Assessing the suitable distribution area of Pinus koraiensis based on an optimized MaxEnt model[J]. Chin J Ecol, 2019, 38(8): 2570-2576. DOI:10.13292/j.1000-4890.201908.017 |
| [10] |
管强, 刘吉平, 武海涛, 等. 中国自然湿地螺类生态学研究进展[J]. 生态学报, 2016, 36(9): 2471-2481. Guan Q, Liu JP, Wu HT, et al. Research progress on the ecology of natural wetland snails (Mollusca: Gastropoda) in China[J]. Acta Ecol Sin, 2016, 36(9): 2471-2481. DOI:10.5846/stxb201411162267 |
| [11] |
曹正光, 蒋忻坡. 几种环境因子对梨形环棱螺的影响[J]. 上海水产大学学报, 1998, 7(3): 200-205. Cao ZG, Jiang XP. The influence of environmental factors on Bellamya purificate[J]. J Shanghai Fish Univ, 1998, 7(3): 200-205. |
| [12] |
李宽意, 刘正文, 杨宏伟, 等. 牧食损害对伊乐藻(Elodea nuttallii)生长的影响[J]. 生态学报, 2007, 27(10): 4209-4213. Li KY, Liu ZW, Yang HW, et al. Effects of mechanical damage on the growth of Elodea nuttallii[J]. Acta Ecol Sin, 2007, 27(10): 4209-4213. DOI:10.3321/j.issn:1000-0933.2007.10.031 |
| [13] |
池秀莲, 唐志尧. 面积、温度及分布区限制对物种丰富度海拔格局的影响: 以秦岭太白山为例[J]. 植物生态学报, 2011, 35(4): 362-370. Chi XL, Tang ZY. Effects of area, temperature and geometric constraints on elevational patterns of species richness: A case study in the Mountain Taibai, Qinling Mountains, China[J]. Chin J Plant Ecol, 2011, 35(4): 362-370. DOI:10.3724/SP.J.1258.2011.00362 |
| [14] |
刘意立. 我国亚热带季风气候区湿地土壤CO2、CH4排放规律研究[D]. 杭州: 浙江大学, 2014. Liu YL. Laboratory study on the carbon dioxide and methane emission from wetland soil in subtropical monsoon climate zone[D]. Hangzhou: Zhejiang University, 2014. (in Chinese) |
| [15] |
刘帅磊, 王赛, 崔永德, 等. 亚热带城市河流底栖动物完整性评价: 以流溪河为例[J]. 生态学报, 2018, 38(1): 342-357. Liu SL, Wang S, Cui YD, et al. Ecological assessment of a subtropical urban river based on the Benthic-Index of Biotic Integrity-Liuxi River[J]. Acta Ecol Sin, 2018, 38(1): 342-357. DOI:10.5846/stxb201612242660 |
| [16] |
刘艳斌, 韩微, 贤振华. 温度对福寿螺生长发育及摄食的影响[J]. 南方农业学报, 2011, 42(8): 901-905. Liu YB, Han W, Xian ZH. Effect of different temperatures on growth, development and feeding of Pomacea canaliculata[J]. J South Agric, 2011, 42(8): 901-905. DOI:10.3969/J.issn.2095-1191.2011.08.018 |
| [17] |
杨海艳. 我国人居适宜性的海拔高度分级研究[D]. 南京: 南京师范大学, 2013. Yang HY. Study on altitude classification of human settlement suitability in China[D]. Nanjing: Nanjing Normal University, 2013. (in Chinese) |
| [18] |
曹福祥, 徐庆军, 曹受金, 等. 全球变暖对物种分布的影响研究进展[J]. 中南林业科技大学学报, 2008, 28(6): 86-89. Cao FX, Xu QJ, Cao SJ, et al. Advances of global warming impact on species distribution[J]. J Cent South Univ For Technol, 2008, 28(6): 86-89. DOI:10.3969/j.issn.1673-923X.2008.06.019 |
| [19] |
姜江, 姜大膀, 林一骅. RCP4.5情景下中国季风区及降水变化预估[J]. 大气科学, 2015, 39(5): 901-910. Jiang J, Jiang DB, Lin YH. Projection of monsoon area and precipitation in China under the RCP4.5 scenario[J]. Chin J Atmos Sci, 2015, 39(5): 901-910. DOI:10.3878/j.issn.1006-9895.1411.14216 |
| [20] |
刘燕. 湖北钉螺生存环境主要影响因素探析[J]. 九江学院学报(自然科学版), 2013, 28(3): 1-4. Liu Y. Main environmental factors affecting Oncomelania snail ecology[J]. J Jiujiang Univ(Nat Sci), 2013, 28(3): 1-4. DOI:10.19717/j.cnki.jjun.2013.03.001 |
| [21] |
赵国平. 中国蜱类空间分布及其危害预测[D]. 北京: 军事科学院, 2018. Zhao GP. Spatial distribution of ticks and their hazard prediction in China[D]. Beijing: Academy of Military Sciences, 2018. (in Chinese) |
| [22] |
李博, 周天军. 基于IPCC A1B情景的中国未来气候变化预估: 多模式集合结果及其不确定性[J]. 气候变化研究进展, 2010, 6(4): 270-276. Li B, Zhou TJ. Projected climate change over China under SRES A1B scenario: Multi-model ensemble and uncertainties[J]. Adv Climate Change Res, 2010, 6(4): 270-276. |