 2022, Vol. 33
2022, Vol. 33扩展功能
文章信息
- 吴瑜燕, 李天奇, 刘钦梅, 王金娜, 罗明宇, 龚震宇
- WU Yu-yan, LI Tian-qi, LIU Qin-mei, WANG Jin-na, LUO Ming-yu, GONG Zhen-yu
- 泥鳅对白纹伊蚊幼虫控制效果初探
- Effect of Misgurnus anguillicaudatus in controlling the larvae of Aedes albopictus: A preliminary study
- 中国媒介生物学及控制杂志, 2022, 33(4): 453-457
- Chin J Vector Biol & Control, 2022, 33(4): 453-457
- 10.11853/j.issn.1003.8280.2022.04.001
-
文章历史
- 收稿日期: 2022-02-07




蚊隶属于昆虫纲双翅目蚊科,是具有刺吸式口器的昆虫,可以传播包括疟疾、登革热、黄热病、基孔肯雅热、寨卡病毒病、斑氏丝虫病、流行性乙型脑炎等在内的一系列疾病,属于卫生“四害”之一[1-4]。近年来,蚊媒传染病防控形势日趋严峻,以登革热为例,全球登革热病例在过去50年间增加了30倍,全球40%人口面临着罹患登革热的风险[5]。在我国,登革热风险范围一直呈现由南向北,自东往西扩展的趋势。随着气候变化,登革热每年至少威胁我国2.78亿居民健康[5]。
传统的蚊虫控制主要是通过化学杀虫剂杀灭成蚊,化学杀虫剂具有见效快、易操作等优点,在既往的蚊媒控制中发挥了巨大的作用。然而,随着杀虫剂的重复使用、过度使用、无效使用等情况的出现,蚊虫抗药性问题越来越突出。根据2019年浙江省白纹伊蚊(Aedes albopictus)抗药性监测数据,浙江省超过70%的地级市对至少1种化学杀虫剂产生了可疑抗性,义乌市对菊酯类主要产品(氯菊酯、高效氯氰菊酯和溴氰菊酯)均已产生抗性[6]。另外,化学杀虫剂还会污染环境,使人民健康受到巨大威胁,越来越多的研究发现,杀虫剂的使用与儿童癌症发生率及儿童神经系统的发育障碍呈正相关关系[7-9]。因此,研发绿色、环保、科学的蚊虫控制技术,对蚊虫的可持续控制和蚊媒传染病的精准防控至关重要,其中生物防制蚊虫由于其对环境友好,对人体健康风险低等优点而备受青睐。本文选择了本土小型鱼类——泥鳅(Misgurnus anguillicaudatus)作为研究对象,探讨了在实验室环境中泥鳅对浙江省主要蚊媒白纹伊蚊的吞食效果,为蚊虫生物防制提供新的思路。
1 材料与方法 1.1 实验对象来源白纹伊蚊幼虫系浙江省疾病预防控制中心病媒生物防制科饲养的敏感品系。泥鳅及柳条鱼(Gambusia affinis,又称食蚊鱼)均购自浙江省杭州市某水产繁殖公司。
1.2 实验器材精密人工气候箱(杭州琦盛电子科技有限公司,型号:QRZ-1600B),长、宽、高分别为38、25和13 cm的小型玻璃鱼缸,电子天平(双圈牌,型号:MP200B),1 000 ml量筒,吸蚊器等。
1.3 实验方法 1.3.1 不同龄期泥鳅对白纹伊蚊幼虫的控制效果研究将采购的泥鳅置于脱氯水中,在温度设定为(28±1)℃、相对湿度为60%~70%的人工气候箱内驯养7 d后进行实验。实验时,将泥鳅根据郑闽泉[10]描述的副性征分成未成年组(A)和成年组(B)。未成年组和成年组泥鳅各随机选择12条,对每条泥鳅进行称重,24 h禁食备用。每个小型玻璃鱼缸中装入4 L的脱氯水,放入1条禁食后的泥鳅,构成一个实验对象。以3个实验对象为一组,将未成年与成年泥鳅各分成4组(未成年泥鳅为A1、A2、A3、A4组;成年泥鳅为B1、B2、B3、B4组),放入4龄白纹伊蚊幼虫,其中A1和B1组放入100条/缸,A2和B2组放入200条/缸,A3和B3组放入400条/缸,A4和B4组放入1 000条/缸。幼蚊放入后,观察1、2、4、6、8和24 h每缸幼蚊的存活及化蛹数。以同等体质量的柳条鱼作为对照组,即根据未成年组与成年组泥鳅的平均体质量,选择多条柳条鱼使其分别与未成年组和成年组泥鳅的平均体质量相等,24 h禁食驯养后放入对照鱼缸中作为对照组,未成年组和成年组对照实验各重复3次,未成年泥鳅对照组每缸投入4龄白纹伊蚊幼虫400条(A0组),成年泥鳅对照组每缸投入4龄白纹伊蚊幼虫1 000条(B0组),观察1、2、4、6、8和24 h每缸幼蚊的存活数及化蛹数。
1.3.2 泥鳅对白纹伊蚊幼虫的7 d持效研究随机选择3条驯养后的未成年泥鳅,称重,禁食24 h备用,对照组选择同等体质量的柳条鱼。每个小型玻璃鱼缸中装入4 L的脱氯水,放入1条禁食后的泥鳅或同等体质量的柳条鱼,组成一个实验对象。每个实验对象缸中加入白纹伊蚊4龄幼虫1 000条,24 h后记录每缸中幼蚊的存活及化蛹数并捞出,重新放入1 000条白纹伊蚊4龄幼虫,依此类推,直至第7天,记录实验组和对照组每天幼蚊的存活数及化蛹数。
1.4 统计学分析利用Excel 2010和SPSS 16.0软件对实验数据进行整理和分析,分别计算放入泥鳅后不同时间点的幼蚊存活率,用统计表进行展示。利用广义线性模型(GLMM)分析相同幼蚊密度组间(未成年泥鳅:A3 vs A0;成年泥鳅:B4 vs B0;未成年与成年泥鳅:A1 vs B1,A2 vs B2,A3 vs B3,A4 vs B4)的食蚊数差异(模型关联选择线性分布,“幼蚊存活数”设为因变量,“分组”“重复观测的时间”设为固定效应主效应,每个“泥鳅个体”设为随机效应主效应)。根据不同时间段内泥鳅吞食幼蚊数量分布类型,选择方差分析或者非参数检验比较不同时间段内泥鳅吞食幼蚊数量的差异。P < 0.05为差异有统计学意义。不同时间幼蚊存活率(%)=3组重复实验组存活的幼蚊及蛹数量之和/3组重复实验组投入幼蚊的总数量×100%;24 h单位体质量泥鳅吞食幼蚊数量(条/g)=3组重复实验24 h内吞食的幼蚊数量之和/3组重复实验泥鳅体质量之和。
2 结果 2.1 未成年泥鳅对白纹伊蚊幼虫的吞食效果未成年组泥鳅体质量为3.9~6.3 g/条[均数±标准差:(4.9±0.69)g/条],匹配的对照组柳条鱼体质量为4.9 g/缸。不同时间段内,各实验组泥鳅吞食白纹伊蚊幼虫效果如表 1所示,A1~A3组幼蚊24 h存活率均为0,A4组幼蚊24 h存活率为1.53%,即日吞食幼蚊数约为984条/(鱼∙d)。配比体质量后的对照组柳条鱼缸内幼虫24 h存活率为3.75%。使用GLMM模型比较幼蚊密度相同(400条/缸)的A3组与对照组吞食幼蚊数量,结果发现A3组泥鳅吞食幼蚊的数量显著高于对照组,差异有统计学意义(F=58.051,P < 0.001)。比较各观察时间段内(0~、1~、2~、4~、6~、8~24 h)泥鳅吞食幼蚊数,结果发现不同时间段泥鳅吞食幼蚊的数量差异无统计学意义(Kruskal-Wallis检验,χ2=3.374,P=0.643)。
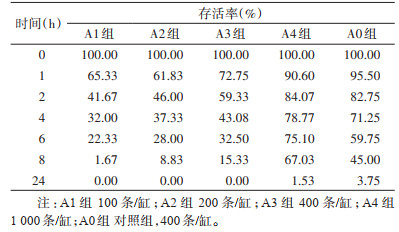
|
成年组泥鳅体质量为16.8~25.5 g/条[均数±标准差:(20.9±2.24)g/条],匹配的对照组柳条鱼体质量为20.9 g/缸。不同时间段内,各成年组泥鳅吞食白纹伊蚊幼虫效果如表 2所示,与未成年组泥鳅吞食幼蚊能力相似,B1~B3组幼蚊24 h存活率均为0,B4组幼蚊24 h存活率为35.23%,即日吞食幼蚊数约为647条/(鱼∙d)。配比体质量后的对照组柳条鱼盆内幼虫24 h存活率为0。使用GLMM模型比较幼蚊密度相同(1 000条/缸)的B4组与对照组吞食幼蚊数量,结果发现B4组泥鳅吞食幼蚊数量显著低于对照组,差异有统计学意义(F=23.162,P < 0.001)。比较各观察时间段内(0~、1~、2~、4~、6~、8~24 h)泥鳅吞食幼蚊数,结果发现不同时间段泥鳅吞食幼蚊的数量差异无统计学意义(Kruskal-Wallis检验,χ2=5.880,P > 0.318)。

|
使用GLMM模型比较相同幼蚊密度组间(A1与B1、A2与B2、A3与B3和A4与B4)成年组与未成年组泥鳅吞食幼蚊数量,结果发现除A3与B3组(400条/缸)外,其余幼蚊密度组中成年组与未成年组泥鳅吞食幼蚊能力差异无统计学意义(100条/缸组:F=0.016,P=0.901;200条/缸组:F=2.238,P=0.143;400条/缸组:F=9.406,P=0.004;1 000条/缸组:F=0.001,P=0.994)。未成年组泥鳅的24 h单位体质量吞食幼蚊数量分别为:20.55条/g(A1),43.48条/g(A2),72.73条/g(A3)和212.52条/g(A4),对照组为78.57条/g;成年组泥鳅的24 h单位体质量泥鳅吞食幼蚊数量分别为:4.86条/g(B1),9.55条/g(B2),18.81条/g(B3)和31.09条/g(B4),对照组为47.85条/g。
2.4 未成年泥鳅7 d连续吞食白纹伊蚊幼虫效果分析3条未成年泥鳅的平均体质量为3.4 g/条,匹配相同体质量的柳条鱼作为对照组,开展7 d连续吞食幼蚊实验。结果发现,实验组第1~7天幼蚊的24 h存活率分别为2.90%、0.73%、2.77%、8.37%、22.27%、15.03%和13.07%。对照组第1~7天幼蚊的24 h存活率分别为65.63%、70.30%、71.67%、71.00%、76.03%、77.40%和72.33%。不同泥鳅间每日吞食白纹伊蚊幼虫数量差异无统计学意义(Kruskal-Wallis检验,χ2=2.650,P=0.266),但实验组每日吞食白纹伊蚊幼虫数量显著高于同等体质量的对照组,差异有统计学意义(Kruskal-Wallis检验,χ2=16.758,P=0.001)。见图 1。

|
| 图 1 未成年组泥鳅连续7 d吞食白纹伊蚊幼虫数量 Figure 1 Number of Aedes albopictus larvae devoured by juvenile Misgurnus anguillicaudatus in 7 consecutive days |
| |
本研究选择了本地小型鱼种——泥鳅作为实验对象,研究其在实验室环境中吞食白纹伊蚊幼虫的效果,以期为我国蚊虫的生物防制提供新的思路。结果发现,在相同白纹伊蚊幼虫密度下,同等体质量的未成年组泥鳅吞食幼蚊的效果优于柳条鱼,每条未成年泥鳅24 h吞食白纹伊蚊幼虫数可达984条;而同等体质量的成年组泥鳅吞食幼蚊的效果不及柳条鱼,每条成年泥鳅24 h吞食白纹伊蚊幼虫数约为647条。相同白纹伊蚊幼虫密度下(除400条/缸外),未成年组泥鳅和成年组泥鳅吞食幼蚊数差异无统计学意义,但单位体质量的未成年泥鳅24 h吞食幼蚊数量,可为成年泥鳅的6.84倍(212.52条/g vs 31.09条/g)。选择未成年泥鳅做7 d连续吞食幼蚊实验,结果显示,中短期内泥鳅每日吞食幼蚊的水平较为稳定,日吞食幼蚊数量差异无统计学意义。
柳条鱼又名食蚊鱼,是一种吞食幼蚊能力较强的胎鳉科小型鱼类,原产美国,因其嗜食幼蚊而于1927年引入我国。李鸿宝等[11]研究表明,在居民小区放置柳条鱼诱蚊缸可显著降低小区蚊密度。因柳条鱼体型较小,本研究通过配比体质量的方式来比较同等体质量下泥鳅与柳条鱼吞食幼蚊的能力。本研究选择了柳条鱼作为对照组,结果发现,柳条鱼的24 h单位体质量吞食幼蚊数量远低于未成年组泥鳅(78.57条/g vs 212.52条/g),即未成年泥鳅具有更强的吞食白纹伊蚊幼虫的能力。另外,研究还发现,未成年组与成年组泥鳅的日吞食幼蚊数差异无统计学意义,未成年组泥鳅并没有因为体型较小而少吞食幼蚊。本实验中未成年组泥鳅体质量为3.2~6.3 g/条,主要为1龄泥鳅,而成年组泥鳅体质量为16.8~25.5 g/条,主要为3~4龄泥鳅[12]。有研究表明,3龄前泥鳅生长速度较快,此后生长减慢,由此推断,未成年组泥鳅优异的吞食幼蚊能力可能与其特定的生长阶段,需要通过摄入大量的食物来维持快速的生长发育有关[12-13]。
蚊虫防制是我国爱国卫生工作的一项重要内容,也是防制蚊传疾病的有效手段。为解决我国南方地区经常发生因输入病例而导致的农村蚊媒传染病暴发问题,促进农村环境改善和以民宿为代表的农村旅游业发展,浙江省率先开展了“无蚊村”建设试点[14],拟通过科学的手段,绿色、经济、可持续地将蚊虫密度控制在不足为害的水平[15]。然而在“无蚊村”创建过程中,我们发现许多农村地区仍存在卫生意识落后、积水和储水习俗普遍等特点,给“无蚊村”创建带来诸多困难[14]。生物防制蚊虫能以绿色、可持续的方式有效控制蚊虫密度,同时减少农民积水和储水习俗影响,适于在农村地区推广。当前生物防制蚊虫主要有2种方法,一是用微生物制剂灭蚊,如苏云金杆菌、球形芽孢杆菌等,二是利用天敌,常见的防治物种有柳条鱼及四大家鱼等,但柳条鱼为外来物种,四大家鱼适合在湖、塘等大型水体里生存,而居民生活环境周围多为小型水体,不适于此类鱼种生存。基于本土鱼类探寻一种新的适用于居家环境幼蚊防治的方法十分关键。泥鳅在全国各地河川、沟渠、水田、池塘及水库等天然淡水水域中均有分布,是极具经济与药用价值的本土鱼类[16-17]。本研究发现,泥鳅具有良好的吞食白纹伊蚊幼虫的能力,未成年泥鳅由于体型小、生命力顽强、保留性好等优点,非常适宜用于居家外环境小型水体,如废桶、水缸等处饲养除蚊[18]。但本研究仅开展了实验室内泥鳅对白纹伊蚊幼虫吞食效果研究,实际应用过程中还可能存在因水体环境、气候等多种因素影响,泥鳅的实际杀灭幼蚊的效果可能有所不同。因此,浙江省在实验室研究的基础上,积极开展了农村外环境中泥鳅控制幼蚊密度的现场实验,成果有待进一步公布。
利益冲突 无
| [1] |
Kampango A, Abílio AP. The Asian tiger hunts in Maputo city-the first confirmed report of Aedes (Stegomyia) albopictus (Skuse, 1895) in Mozambique[J]. Parasit Vectors, 2016, 9: 76. DOI:10.1186/S13071-016-1361-4 |
| [2] |
龚道方, 周红宁. 中国登革热重要媒介白纹伊蚊的研究进展[J]. 中国媒介生物学及控制杂志, 2009, 20(6): 607-610. Gong DF, Zhou HN. Progress in dengue fever important vector Aedes albopictus in China[J]. Chin J Vetor Biol Control, 2009, 20(6): 607-610. |
| [3] |
Luo QC, Hao YJ, Meng FX, et al. The mitochondrial genomes of Culex tritaeniorhynchus and Cx. pipiens pallens (Diptera: Culicidae) and comparison analysis with two other Culex species[J]. Parasit Vectors, 2016, 9: 406. DOI:10.1186/s13071-016-1694-z |
| [4] |
Rašić G, Schama R, Powell R, et al. Contrasting genetic structure between mitochondrial and nuclear markers in the dengue fever mosquito from Rio de Janeiro: Implications for vector control[J]. Evol Appl, 2015, 8(9): 901-915. DOI:10.1111/eva.12301 |
| [5] |
刘起勇. 新时代媒介生物传染病形势及防控对策[J]. 中国媒介生物学及控制杂志, 2019, 30(1): 1-6, 11. Liu QY. Epidemic profile of vector-borne diseases and vector control strategies in the new era[J]. Chin J Vetor Biol Control, 2019, 30(1): 1-6, 11. DOI:10.11853/j.issn.1003.8280.2019.01.001 |
| [6] |
Hou J, Liu QM, Wang JN, et al. Insecticide resistance of Aedes albopictus in Zhejiang province, China[J]. Biosci Trends, 2020, 14(4): 248-254. DOI:10.5582/bst.2020.03194 |
| [7] |
Alavanja MCR, Hoppin JA, Kamel F. Health effects of chronic pesticide exposure: Cancer and neurotoxicity[J]. Annu Rev Public Health, 2004, 25: 155-197. DOI:10.1146/annurev.publhealth.25.101802.123020 |
| [8] |
Daniels JL, Olshan AF, Savitz DA. Pesticides and childhood cancers[J]. Environ Health Perspect, 1997, 105(10): 1068-1077. DOI:10.1289/ehp.971051068 |
| [9] |
Rosas LG, Eskenazi B. Pesticides and child neurodevelopment[J]. Curr Opin Pediatr, 2008, 20(2): 191-197. DOI:10.1097/MOP.0b013e3282f60a7d |
| [10] |
郑闽泉. 泥鳅繁殖习性的观察[J]. 四川动物, 1992, 11(2): 35-36. Zheng MQ. Observation on the reproductive habits of Misgurnus anguillicaudatus[J]. Sichuan J Zool, 1992, 11(2): 35-36. |
| [11] |
李鸿宝, 徐宏, 张爱凤. 居民小区设置柳条鱼诱蚊缸实验效果观察[J]. 上海预防医学杂志, 2011, 23(2): 75-76. Li HB, Xu H, Zhang AF. Observation on the experimental effect of setting up wicker-fish tank mosquito traps in residential quarters[J]. Shanghai J Prev Med, 2011, 23(2): 75-76. DOI:10.3969/j.issn.1004-9231.2011.02.014 |
| [12] |
王敏, 王卫民, 鄢建龙. 泥鳅和大鳞副泥鳅年龄与生长的比较研究[J]. 水利渔业, 2001, 21(1): 7-9. Wang M, Wang WM, Yan JL. Comparative studies on the age and growth of Misgurnus anguillicaudatus and Paramisgurnus dabryanus[J]. Rreservoir Fisheries, 2001, 21(1): 7-9. DOI:10.3969/j.issn.1003-1278.2001.01.003 |
| [13] |
王坤, 凌去非, 李倩, 等. 苏州地区泥鳅和大鳞副泥鳅年龄与生长的初步研究[J]. 上海海洋大学学报, 2009, 18(5): 553-558. Wang K, Ling QF, Li Q, et al. Primary study on the age and growth of Misgurnus anguillicaudatus and Paramisgurnus dabryanus in the area of Suzhou[J]. J Shanghai Ocean Univ, 2009, 18(5): 553-558. |
| [14] |
陈恩富, 郭颂, 黄文忠, 等. 农村蚊虫防制和"无蚊村"建设评价指标体系[J]. 预防医学, 2019, 31(3): 217-220. Chen EF, Guo S, Huang WZ, et al. The mosquito control and evaluation system of "mosquito-free village" construction in rural areas[J]. Prev Med, 2019, 31(3): 217-220. DOI:10.19485/j.cnki.issn2096-5087.2019.03.001 |
| [15] |
朱海博, 叶慧锋, 陈明, 等. 浙江省青田县"无蚊村"建设的效果评价[J]. 中国媒介生物学及控制杂志, 2021, 32(3): 365-368. Zhu HB, Ye HF, Chen M, et al. Effectiveness evaluation of "mosquito-free village" construction in Qingtian county, Zhejiang province, China[J]. Chin J Vector Biol Control, 2021, 32(3): 365-368. DOI:10.11853/j.issn.1003.8280.2021.03.021 |
| [16] |
张莲姬. 泥鳅鱼黑色素抗氧化作用[J]. 食品研究与开发, 2010, 31(8): 11-14. Zhang LJ. Study on the antioxidant of the loach melanin[J]. Food Res Dev, 2010, 31(8): 11-14. DOI:10.3969/j.issn.1005-6521.2010.08.004 |
| [17] |
钦传光, 陈泽宪, 周军, 等. 泥鳅及其提取物中微量元素的含量分析[J]. 微量元素与健康研究, 2001, 18(1): 49-50. Qin CG, Chen ZX, Zhou J, et al. Analysis of trace elements in the loach, Misgurnus angillicaudatus, and its extractives[J]. Stud Trace Elem Health, 2001, 18(1): 49-50. DOI:10.3969/j.issn.1005-5320.2001.01.025 |
| [18] |
冯大庆. 泥鳅实验杂谈[J]. 生物学通报, 1992, 37(1): 39. Feng DQ. Talking about loach experiment[J]. Bull Biol, 1992, 37(1): 39. |