 2022, Vol. 33
2022, Vol. 33扩展功能
文章信息
- 胡艳红, 王姝懿, 李建云, 张大禹, 冯一兰
- HU Yan-hong, WANG Shu-yi, LI Jian-yun, ZHANG Da-yu, FENG Yi-lan
- 内蒙古自治区2020年鼠疫监测调查与分析
- Analysis of plague surveillance in Inner Mongolia Autonomous Region, China, 2020
- 中国媒介生物学及控制杂志, 2022, 33(3): 418-425
- Chin J Vector Biol & Control, 2022, 33(3): 418-425
- 10.11853/j.issn.1003.8280.2022.03.020
-
文章历史
- 收稿日期: 2021-10-18




内蒙古自治区(内蒙古)地处中国北部,位于北纬37°24′~53°23′,东经97°12′~126°04′,东部与黑龙江、吉林、辽宁和河北省交界,南部与宁夏、陕西、山西省(自治区)相邻,西南部与甘肃省毗邻,北部与俄罗斯、蒙古国接壤,国境线长4 200 km[1-3]。全区鼠疫自然疫源地面积约33.7万hm2,占全区行政区划总面积的1/3,有蒙古旱獭(Marmaota sibibirica)、达乌尔黄鼠(Spermophilus dauricus)、布氏田鼠(Microtus brandti)、长爪沙鼠(Meriones unguiculatus)4种类型鼠疫疫源地[4-6],具有疫源地面积大、类型多、结构复杂的特点,是鼠疫危害较为严重的地区[7-9]。目前,全区有现疫鼠疫流行县(旗、市、区)24个。
2020年,在全区12个盟(市)的57个鼠疫监测县(旗、市、区)设立固定监测点18个(国家级8个、省级10个)、流动监测点109个、鼠情调查点5个、疫源检索点47个。2020年,全区共报告3例人间鼠疫病例,长爪沙鼠、布氏田鼠和达乌尔黄鼠鼠疫疫源地均发生了动物间鼠疫疫情。本文收集内蒙古地区2020年鼠疫监测数据,旨在掌握鼠疫宿主和媒介蚤的种类和数量,深入分析鼠疫流行特征,为制定有效的防控措施提供科学依据。
1 材料与方法 1.1 监测范围2020年1-12月全区12个盟(市)57个监测县(旗、市、区)按照《全国鼠疫监测工作方案》和《内蒙古自治区鼠疫监测工作方案》要求开展鼠疫监测工作,资料来源于各地上报的监测数据。
1.2 宿主调查 1.2.1 鼠密度按照地貌、植被和主要宿主动物数量等指标划分栖息地,按不同栖息地面积的0.5%比例分层抽样,采用路线法、样方法、弓形夹法等进行鼠类密度调查。
1.2.2 夜行鼠根据不同的生境运用5 m夹线法在不同栖息地调查夜行鼠的活动,布夹数根据不同疫源地要求不同,每晚布夹次日早晨收夹。统计夜行鼠的种类和数量。
1.3 媒介蚤采用样方法、逐日捕鼠法、5 m夹线法和收集自毙动物的方法捕获鼠类。所捕鼠需单独装袋,乙醚麻醉后梳检鼠体蚤,在低倍解剖镜下进行分类和计数。
1.4 病原学及血清学检测对收集、捕获的啮齿动物进行包括显微镜涂片检查、细菌培养、噬菌体试验在内的鼠疫细菌学检测,蚤的鼠疫细菌培养,鼠类动物血样的血清学检测采用间接血凝试验(IHA)。实验室检测和阳性的判断以《鼠疫诊断标准》WS 279-2008为依据。
1.5 数据处理采用Excel 2007软件录入全区数据,运用描述性流行病学方法统计分析数据。
2 结果 2.1 人间鼠疫监测2020年,全区报告3例人间鼠疫病例,分别来自巴彦淖尔市乌拉特中旗、乌拉特前旗和包头市达茂旗。其中腺鼠疫病例2例,肠鼠疫1例。
2.2 动物间鼠疫监测在6个盟(市)的16个县(旗、市、区)共检出动物鼠疫耶尔森菌(Yersinia pestis,鼠疫菌)99株、阳性血清6份、判定疫点51个。在99株鼠疫菌中,从疫鼠分离到的鼠疫菌为84株,从疫蚤分离到的鼠疫菌为15株(疫蚤共52匹分15组,每组均分离到1株鼠疫菌)(表 1)。长爪沙鼠鼠疫疫源地的乌兰察布市检出鼠疫菌最多,4个旗(县)共检出31株,占31.31%;其次是包头市的达茂旗检出鼠疫菌22株,占22.22%;巴彦淖尔市共在2个旗(县)检出鼠疫菌15株,占15.15%。在长爪沙鼠、布氏田鼠和达乌尔黄鼠鼠疫疫源地发现了动物间鼠疫疫情。
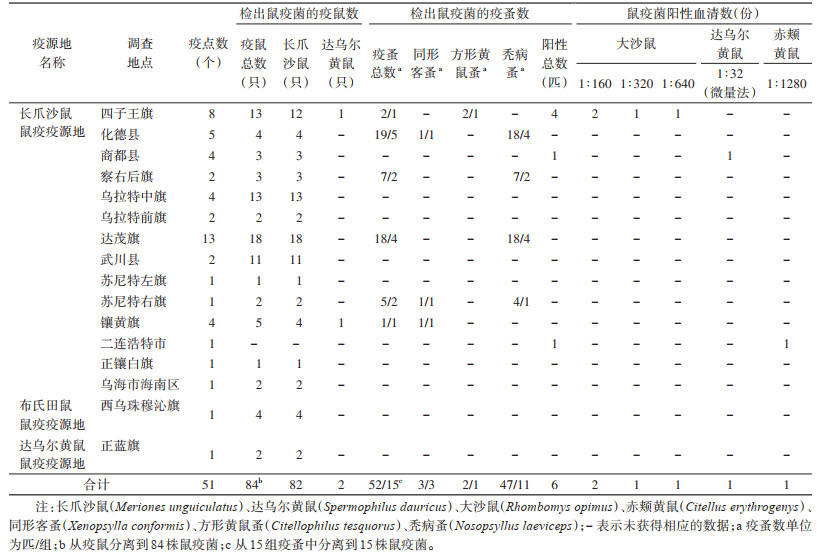
|
采用病原学分离培养方法检测宿主动物22 451只,检出阳性材料84份,阳性率为0.37%,分离出鼠疫菌84株,其中长爪沙鼠的阳性菌株82份,占97.62%,来自达乌尔黄鼠的阳性菌株2份,占2.38%;蚤类检测21 821匹,分成7 196组,检出阳性材料52份,阳性率为0.24%,分离出鼠疫菌15株。
采用IHA对10 671份血清进行检测,其中达乌尔黄鼠、长爪沙鼠和子午沙鼠(M. meridianus)的血清各检测8 761、1 118和465份,分别占总数的82.10%、10.48%和4.36%。分离阳性血清6份,阳性率为0.06%。6份阳性血清中有4份来自大沙鼠(Rhombomys opimus),1份来自达乌尔黄鼠,另1份来自赤颊黄鼠(Citellus erythrogenys)。
2.3 宿主动物调查结果 2.3.1 主要宿主密度及主要鼠种内蒙古全年累计监测面积共计123 848.5 km2,约占全区疫源地面积的36.83%。全区对宿主动物密度调查共计6 003 hm2,其中采用样方法调查5 133 hm2,路线法调查870 hm2。捕获各种宿主动物共计22 710只,其中主要宿主动物7 617只。主要宿主动物以长爪沙鼠为主,捕获6 059只,占79.55%;达乌尔黄鼠次之,捕获1 167只,占15.32%;布氏田鼠捕获153只,占2.00%。鄂尔多斯市鼠密度最高,为4.88只/hm2,赤峰市最低,为0.06只/hm2(表 2)。鼠密度最高的旗(县)为长爪沙鼠鼠疫疫源地鄂尔多斯高原的鄂托克前旗,其次是长爪沙鼠鼠疫疫源地乌兰察布高原的武川县,分别为7.41和4.24只/hm2。
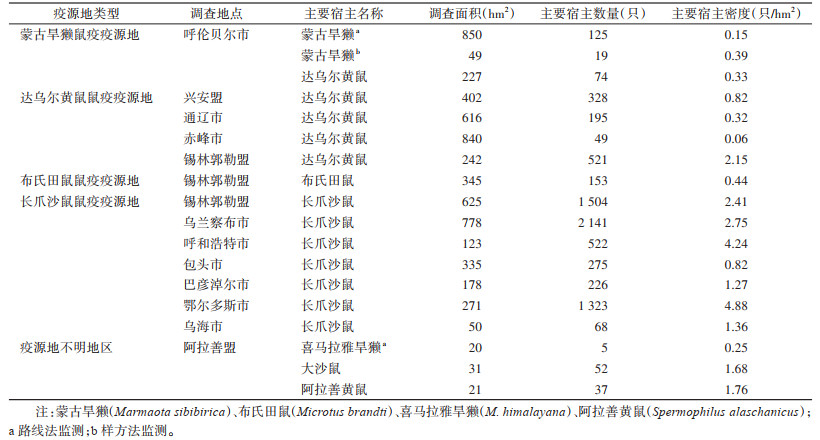
|
共布夹81 560次,捕鼠19种1 833只,捕获率为2.25%。其中黑线仓鼠(Cricetulus barabensis)462只,占25.20%,子午沙鼠(M. meridianus)324只,占17.68%,五趾跳鼠(Allactaga sibirica)289只,占15.77%,以上3种鼠为该地区的主要夜行鼠,也是优势鼠种。长耳跳鼠(Euchoreutes naso)、短耳仓鼠(C. eversmanni)、黑线姬鼠(Apodemus agrarius)和达乌尔鼠兔(Ochotona dauurica)为稀有鼠种,其他常见鼠种见表 3。蒙古旱獭鼠疫疫源地夜间捕获的鼠以长爪沙鼠为主,占该疫源地鼠种的39.20%(49/125)。达乌尔黄鼠鼠疫疫源地松辽平原和察哈尔丘陵地区夜行鼠均以黑线仓鼠为主,分别占61.53%(379/616)和66.67%(24/36)。布氏田鼠鼠疫疫源地夜行鼠以五趾跳鼠为优势鼠种,占54.76%(23/43)。长爪沙鼠鼠疫疫源地乌兰察布高原和鄂尔多斯高原地区夜间捕获的鼠类分别以子午沙鼠和长爪沙鼠为主,分别占30.43%(140/460)和38.65%(160/414)。疫源地不明的阿拉善盟地区以子午沙鼠[70.00%(98/140)]为主。

|
共梳检鼠类13143只,染蚤宿主数3 993只,捕获寄生蚤12 798匹,染蚤率为30.38%,总蚤指数为0.97(表 4)。蒙古旱獭鼠疫疫源地、达乌尔黄鼠鼠疫疫源地松辽平原和察哈尔丘陵地区均以方形黄鼠蚤为主要蚤种,其所占该疫源地蚤类比例分别为99.61%(511/513)、94.69%(2 499/2 639)和79.19%(2 611/3 297);布氏田鼠鼠疫疫源地近代新蚤(Neopsylla pleskei)为优势蚤种,占比为37.44%(73/195);长爪沙鼠鼠疫疫源地乌兰察布高原和鄂尔多斯高原地区以秃病蚤(1 540/3 182,48.40%)和同形客蚤(1 526/2 919,52.28%)为主;疫源地不明的阿拉善盟地区以阿巴盖新蚤(N. abagaitui)(31/53,58.49%)为主。

|
全区监测点共探洞8 184个,染蚤洞808个,染蚤总数1 987匹,染蚤率为9.87%,蚤指数为0.24(表 5)。以方形黄鼠蚤为优势蚤种,占获蚤总数的89.03%(1 769/1 987);共挖巢829个,染蚤巢数172个,获蚤总数2 005匹,染蚤率为20.75%,蚤指数为2.42。其中方形黄鼠蚤1 152匹,占比最高(57.46%)。见表 6。
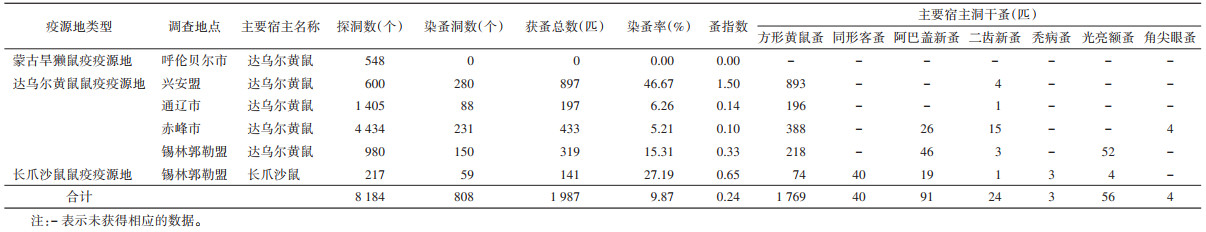
|
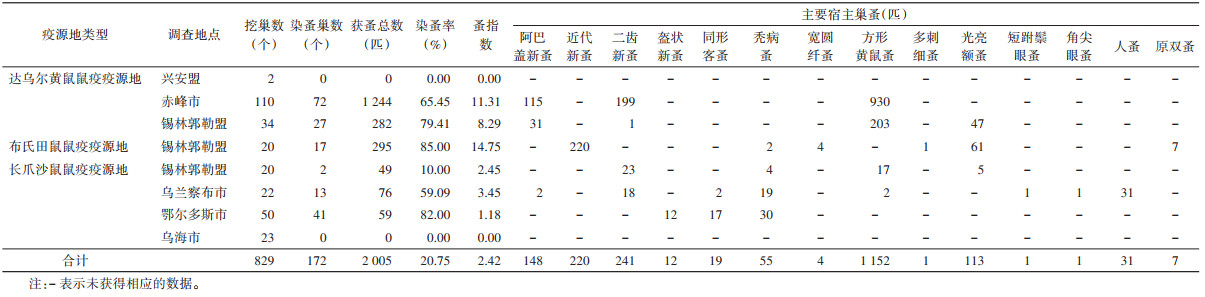
|
内蒙古地区2019年报告人间鼠疫2例,2020年3例,人间鼠疫病例均生活在鼠疫历史疫源地区,职业均为牧民,在其牧场放牧时感染发病。人间鼠疫疫情发生后,自治区加强组织领导,成立党委政府领导的指挥组,强化医疗救治,划定隔离封锁范围,加强公众鼠疫防治知识宣传教育工作。大力开展爱国卫生运动,加强鼠防防控工作经费及物资保障。通过群防群控、联防联控等强有力的措施,取得了无继发病例出现、疫情未扩散的成绩。
3.2 动物间鼠疫近年来,内蒙古地区动物间鼠疫疫情活跃,流行强度大、涉及地区多、流行范围广。在达乌尔黄鼠、布氏田鼠和长爪沙鼠3型鼠疫疫源地连续发生多起以长爪沙鼠为主的动物间鼠疫疫情,部分县(旗、市、区)连年发生动物鼠疫,还有个别县(旗、市、区)时隔多年再次发生疫情。在长爪沙鼠鼠疫疫源地动物鼠疫出现了多点暴发流行,在整个疫源地近14万km2范围内呈不断扩大态势。在达乌尔黄鼠鼠疫疫源地的正蓝旗、布氏田鼠鼠疫疫源地的西乌珠穆沁旗以及自建市以来从未发生过动物间鼠疫的乌海市海南区,均发生了动物间鼠疫疫情,流行范围之广为历年之最,且疫情逐渐向人群聚集区靠近。动物疫情在乌拉特中旗、达茂旗等南部局部地区的半农半牧或接近农区的草原地带仍呈扩大蔓延态势,出现了由既往动物鼠疫频发的草原向半农半牧区扩散的趋势,对人类构成了严重威胁。
2020年达乌尔黄鼠鼠疫疫源地鼠密度均有所下降,兴安盟主要宿主达乌尔黄鼠密度为0.82只/hm2,从近3年监测数据来看(2017年为1.47只/ hm2、2018年为1.29只/ hm2、2019年为0.92只/ hm2),鼠密度整体呈现连年下降的趋势,其原因可能是近几年该地区生态恢复措施得到落实,效果明显,植被种类繁多且生长旺盛,其环境不适合黄鼠生存。通辽市主要宿主达乌尔黄鼠密度为0.32只/hm2,从近3年监测数据来看(2017年为0.58只/hm2、2018年为0.46只/hm2、2019年为0.38只/hm2),鼠密度整体呈现逐年下降的趋势。分析原因,可能是由于近几年农、牧、林、水业的快速发展,人为改变疫源地的原有生态环境因素增多,使黄鼠的栖息环境发生明显变化,生存环境不断缩小,群落之间的相互迁徙受到限制。
长爪沙鼠鼠疫疫源地近年来动物间疫情持续发生,2019和2020年接连发生人间鼠疫,主要宿主和媒介数量多、分布广,而且疫点、疫区有向城镇、人口密集区逼近的趋势,对周围群众构成了极大威胁。长爪沙鼠密度除了呼和浩特市武川县、巴彦淖尔市部分地区以外,其他地区相对有些许下降,其原因一是因为春季疫情流行直接导致鼠密度下降,二是由于各县(旗、市、区)均开展了大范围的灭鼠工作。通过对动物间鼠疫踏查、监测及流行情况进行综合分析研判,动物间鼠疫流行已逐渐由峰值期转入下滑期,但是由于疫源地面积大,动物间鼠疫发生的风险依然存在,同时不能排除人间散发病例的可能性,疫情防控形势仍然严峻[10-12]。由于生境的变化,长爪沙鼠分布呈现扩大的趋势,在未踏查的地区不排除存在动物间鼠疫流行的可能性[13-15],各地区均面临传入人间病例的可能性,但扩散蔓延的可能性较低。综合得出全区长爪沙鼠鼠疫疫源地鼠密度均较高,应加强该地区的监测工作。
3.3 鼠疫防控建议根据近年的鼠疫疫源地监测情况,锡林郭勒盟、赤峰市、乌兰察布市等几个环北京地区应提高警惕。内蒙古鼠疫防控形势依然严峻,动物间鼠疫传入人间的风险依然存在。在今后的工作中,应继续采取各项综合性的防控措施,加大监测工作力度,扩大疫源地检索范围,及时发现动物疫情,并有效处置疫情,全面构筑“鼠传人”“人传人”“向外扩散”三道防线,坚决降低人间疫情发生和传播的风险。
利益冲突 无
| [1] |
段天一. 1997-2016年内蒙古自治区长爪沙鼠鼠疫自然疫源地鼠疫监测流行病特征分析[D]. 长春: 吉林大学, 2018. Duan TY. Analysis on epidemic characteristics of plague monitoring in natural foci of Mongolian gerbil in Inner Mongolia Autonomous Region from 1997 to 2016[D]. Changchun: Jilin University, 2018. (in Chinese) |
| [2] |
张芳, 李昊, 田洁, 等. 内蒙古高原长爪沙鼠鼠疫自然疫源地现状分析[J]. 中国地方病防治, 2020, 35(3): 220-222. Zhang F, Li H, Tian J, et al. Analysis on the current situation of the natural foci of gerbil plague in Inner Mongolia plateau[J]. Chin J Ctrl Endem Dis, 2020, 35(3): 220-222. |
| [3] |
李建云, 张忠兵, 范蒙光, 等. 内蒙古长爪沙鼠鼠疫疫源地现状分析[J]. 医学动物防制, 2015, 31(7): 722-724,727. Li JY, Zhang ZB, Fan MG, et al. Meriones unguiculatus plague foci status analysis of Inner Mongolia[J]. J Med Pest Control, 2015, 31(7): 722-724,727. DOI:10.7629/yxdwfz201507005 |
| [4] |
杜国义, 杨建明, 王海峰, 等. 中国鼠疫自然疫源地宿主多样性研究进展[J]. 中国媒介生物学及控制杂志, 2012, 23(3): 273-274. Du GY, Yang JM, Wang HF, et al. Advances in research on the host diversity in plague natural foci in China[J]. Chin J Vector Biol Control, 2012, 23(3): 273-274. |
| [5] |
Liu W, Wan X, Zhong W. Population dynamics of the Mongolian gerbils: Seasonal patterns and interactions among density, reproduction and climate[J]. J Arid Environ, 2007, 68(3): 383-397. DOI:10.1016/j.jaridenv.2006.07.002 |
| [6] |
龚正达, 于心, 刘起勇, 等. 中国鼠疫自然疫源地分型研究Ⅵ. 鼠疫媒介生物学特征[J]. 中华流行病学杂志, 2012, 33(8): 818-822. Gong ZD, Yu X, Liu QY, et al. Ecological-geographic landscapes of natural plague foci in China Ⅵ. Biological characteristics of natural vectors of Yesinia pestis[J]. Chin J Epidemiol, 2012, 33(8): 818-822. DOI:10.3760/cma.j.issn.0254-6450.2012.08.014 |
| [7] |
丛显斌, 刘振才. 中国鼠疫及其防治(2001-2010)[M]. 长春: 吉林科学技术出版社, 2014: 5-13. Cong XB, Liu ZC. Chinese plague and its control (2001-2010)[M]. Changchun: Jilin Science and Technology Press, 2014: 5-13. |
| [8] |
李保荣, 石杲, 刘俊. 内蒙古高原长爪沙鼠鼠疫疫源地媒介现状分析[J]. 中国媒介生物学及控制杂志, 2013, 24(3): 249-251. Li BR, Shi G, Liu J. Analysis of current situation of plague vectors in Meriones unguiculatus plague foci on Inner Mongolia plateau[J]. Chin J Vector Biol Control, 2013, 24(3): 249-251. |
| [9] |
许磊, 方喜业, 周冬生, 等. 中国鼠疫自然疫源地的演化动态及环境生态位的生物学特征[J]. 中国媒介生物学及控制杂志, 2015, 26(3): 228-232. Xu L, Fang XY, Zhou DS, et al. The evolutionary dynamics and the ecological niche of natural plague foci in China[J]. Chin J Vector Biol Control, 2015, 26(3): 228-232. DOI:10.11853/j.issn.1003.4692.2015.03.003 |
| [10] |
尉瑞平, 范蒙光, 李建云, 等. 内蒙古长爪沙鼠鼠疫疫源地蚤类的某些特点[J]. 中国媒介生物学及控制杂志, 2014, 25(2): 170-173. Yu RP, Fan MG, Li JY, et al. Some characteristics of flea populations in plague natural foci of Meriones unguiculatus in Inner Mongolia, China[J]. Chin J Vector Biol Control, 2014, 25(2): 170-173. DOI:10.11853/j.issn.1003.4692.2014.02.023 |
| [11] |
毛乌力吉. 锡林郭勒盟长爪沙鼠鼠疫疫源地动物鼠疫流行相关因素分析[J]. 中国地方病防治杂志, 2017, 32(4): 391. Mao WLJ. Analysis of related factors of animal plague epidemic in the gerbil plague foci in XilinGol League[J]. Chin J Ctrl Endem Dis, 2017, 32(4): 391. |
| [12] |
Pan Q, Li M, Shi YL, et al. Lipidomics reveals mitochondrial membrane remodeling associated with acute thermoregulation in a rodent with a wide thermoneutral zone[J]. Lipids, 2014, 49(7): 715-730. DOI:10.1007/s11745-014-3900-0 |
| [13] |
张学英, 迟庆生, 刘伟, 等. 长爪沙鼠的行为和生理生态学研究进展[J]. 中国科学: 生命科学, 2016, 46(1): 120-128. Zhang XY, Chi QS, Liu W, et al. Studies of behavioral and physiological ecology in Mongolian gerbils (Meriones unguiculatus)[J]. Sci Sin Vitae, 2016, 46(1): 120-128. DOI:10.1360/N052015-00223 |
| [14] |
赵秋芳, 尹家祥. 鼠疫自然疫源地构成成分及其作用分析[J]. 现代预防医学, 2016, 43(2): 370-372. Zhao QF, Yin JX. Analysis for the compositions of plague natural foci and their roles[J]. Mod Prev Med, 2016, 43(2): 370-372. |
| [15] |
Shi YL, Chi QS, Liu W, et al. Environmental metabolomics reveal geographic variation in aerobic metabolism and metabolic substrates in Mongolian gerbils (Meriones unguiculatus)[J]. Comp Biochem Physiol Part D: Genomics Proteomics, 2015, 14: 42-52. DOI:10.1016/j.cbd.2015.03.001 |