 2022, Vol. 33
2022, Vol. 33扩展功能
文章信息
- 刘郁桐, 刘雨杭, 袁道欢, 王宇杰, 秦姣, 王健, 陈鸿辉, 王英永, 陈再雄, 聂海燕, 刘全生
- LIU Yu-tong, LIU Yu-hang, YUAN Dao-huan, WANG Yu-jie, QIN Jiao, WANG Jian, CHEN Hong-hui, WANG Ying-yong, CHEN Zai-xiong, NIE Hai-yan, LIU Quan-sheng
- 广东韶关丹霞山国家级自然保护区内鼠种构成及分布调查分析
- Species composition and distribution of rodents in Danxia Mountain National Nature Reserve, Shaoguan, Guangdong province, China
- 中国媒介生物学及控制杂志, 2022, 33(1): 94-99
- Chin J Vector Biol & Control, 2022, 33(1): 94-99
- 10.11853/j.issn.1003.8280.2022.01.017
-
文章历史
- 收稿日期: 2021-08-26




2 广东省科学院动物研究所, 广东省动物保护与资源利用重点实验室, 广东省野生动物保护与利用公共实验室, 广东 广州 510260;
3 中山大学, 生命科学学院, 广东 广州 510275;
4 韶关市丹霞山管理委员会, 广东 韶关 512300
2 Guangdong Key Laboratory of Animal Conservation and Resource Utilization, Guangdong Public Laboratory of Wild Animal Conservation and Utilization, Institute of Zoology, Guangdong Academy of Sciences, Guangzhou, Guangdong 510260, China;
3 School of Life Science, Sun Yat-sen University, Guangzhou, Guangdong 510275, China;
4 Administrative Commission of Danxiashan National Park, Shaoguan, Guangdong 512300, China
啮齿动物是哺乳动物中种类最多、分化最大的类群[1]。截至2021年6月底,中国记录到哺乳动物12目59科254属686种,其中啮齿目12科85属235种[2],广东省记录到啮齿目6科17属35种[3]。至2021年我国啮齿目动物较2017年又新增15个新种[2, 4]。啮齿动物由于自身的生理、生态特点,对生境变化较为敏感,常常被用作生态环境的指示物种[5]。啮齿动物群落结构可反映生境的植物盖度、资源丰富度、动物对资源的利用方式及种间竞争关系等[6]。在一定范围的生态系统中,啮齿动物作为消费者,也是物质和能量的传递者。因其个体小、代谢快、分布广,一定程度上加速了物质循环及能量流动速率。但受限于个体小,能获得的生存资源有限,因而啮齿类对栖息地环境、植被组成更为依赖。因此,不同生境中啮齿动物的种类、数量、种群结构均存在差异[6-7]。一些啮齿动物储存种子的习性促进植物群落的更新和演替。
随着经济快速发展,人类活动使环境异质性增加,这提高了鼠类的物种丰富度及繁殖成功率。而其数量过多则会形成“鼠害”,影响农、林、牧等行业的生产。此外,啮齿动物可贮存、传播多种人兽共患病,威胁着人类生命健康。而随着旅游业的发展,人类在景区活动激增也使得与野生鼠类的接触机会加大。因此,对景区内啮齿动物种类及分布的调查,对该区域生物多样性保护和鼠害控制均有重要意义。
广东韶关丹霞山国家级自然保护区(丹霞山保护区)地处南岭山脉中段南麓,植被分布广泛,除大面积分布常绿阔叶林外,还有华南亚热带雨林[8],于1995年被认定为国家级自然保护区。因其地质独特、风景秀丽,于2001年被评为国家5A级景区,旅游业兴旺。据韶关市丹霞山管理委员会统计,2019年景区接待旅游人数高达290.5万人次。保护区内及周边地势低平区域散布有许多村落及农田,农业生产活动频繁,但常年鼠害严重。迄今为止,丹霞山保护区内鼠类的系统调查十分缺少,尚不清楚该区域农林主要害鼠种类及分布,盲目沿用其他地区的防治手段,往往使防治工作事倍功半。为此,2019年8月我们以夹捕法在保护区不同生境进行了鼠种类、相对数量及分布的野外调查,并采用形态学和DNA条码技术对捕获鼠类进行鉴定,分析比较农区、景区和林区鼠类种群数量及组成多样性的差异。研究结果可为保护区农林鼠害防治及游客和居民的健康防护提供科学依据。
1 材料与方法 1.1 研究地概况丹霞山保护区位于广东省韶关市仁化县和浈江区境内,面积292 km2,地理坐标113°36′25″~113°47′53″E,24°51′48″~25°04′12″N;年平均气温19.7 ℃,1月平均气温9.3 ℃,7月平均气温28.4 ℃,年降雨量1 715 mm[8]。
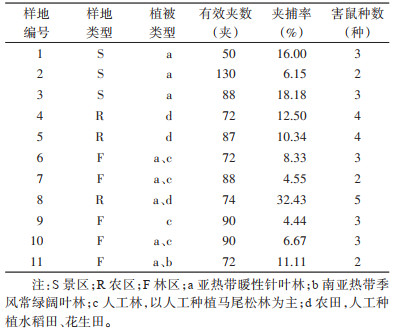
1.2 野外调查方法 1.2.1 样地选择因本次调查目的在于了解人类活动频繁区域鼠类组成,采样范围根据道路可通行情况分散于整个保护区,主要沿仁化河西侧非核心区布点。按照不同生境类型相隔1 km以上选取一定范围片区作为一个样点,共11个样点(图 1)。11个大样点按生境类型分为3类:景区(S,有景点分布,日游客量较大,但居民房屋较少,布夹于景点周围树林内)、农区(R,选取远离景点的居民聚集地,布夹于农田及周围)、林区(F,选取远离景区的丹霞山保护区内部,即未经旅游开发的林区),各区设置3~5个小样点,每个小样点布夹≥50夹。

|
| 注:1~11样地编号。 图 1 广东韶关丹霞山国家级自然保护区调查样地分布 Figure 1 Distribution of sampling sites in Danxia Mountain National Nature Reserve, Shaoguan, Guangdong province |
| |
调查于2019年8月进行。采用目前国内最常见的夹捕法,此种方法适于地栖小型啮齿动物的调查,便于取样、测量身体数据、标本制作及后期分子生物学鉴定。每个样点设置3~5条样线,样线间距50 m以上,夹距5~10 m,每条样线放15~30夹。于16:00-18:30以带壳熟花生作饵放置于捕鼠夹,次日07:00-09:00取回捕鼠夹及被捕获动物。记录具体的样点信息,如植被组成、经纬度等信息。对捕获动物体表杀虫清理后,记录初判种类、性别、体质量(精确到0.1 g)后,测量外部形态参数(mm),保留完整个体的皮张和头骨,留做标本,取后腿肌肉或尾部存于无水乙醇中用于后续分子生物学测定。
1.3 样品形态测量和分子生物学检测 1.3.1 形态学测量记录捕获鼠体各部位(如身体背部、腹部、侧面、尾部及足背)毛色、毛种(是否具针毛)及特殊特征,测量其体质量、头体长、尾长、后足长、耳高等形态参数,初步确定科、属、种[9-10]。
1.3.2 DNA提取、PCR扩增与序列测定使用Universal Genomic DNA Kit(北京康为世纪生物科技有限公司)对肌肉组织进行总DNA提取,保存于-20 ℃备用。使用线粒体细胞色素C氧化酶亚基Ⅰ(COⅠ)通用引物BatL5310(CCTACTCRGCCATTTTACCTATG)和R6036R(ACTTCTGGGTGTCCAAAGAATCA)对所提基因片段进行PCR扩增[11-12]。反应体系为20 μl,2×Es Taq MasterMix(Dye)(北京康为世纪生物科技有限公司)10 μl,正、反向引物各1 μl,模板DNA 1 μl,ddH2O 7 μl。反应条件:95 ℃预变性3 min;95 ℃变性1 min,53 ℃退火30 s,72 ℃延伸1 min,30个循环;72 ℃终延伸5 min。扩增产物送广州天一辉远基因科技有限公司进行单向测序。
1.4 数据处理与分析物种鉴定:使用美国国立生物技术信息中心(NCBI,www.ncbi.nlm.nih.gov)网站BLAST工具进行序列比对,在GenBank中搜索测序结果的相似序列,参照文献[11-17],下载相关物种的序列,使用MEGA 7.0软件以最大似然法(maximum-likelihood,ML)建树,计算遗传距离,对比后确定具体物种。
有效夹捕率(鼠密度):确定样本物种及数量,剔除捕获到非鼠类动物的鼠夹后,计算不同生境的夹捕率。

|
分鼠种夹捕率:确定不同生境捕获到每种鼠类的数量,计算分鼠种夹捕率。
生态学指数:(1)丰富度指数[18](R)=(S-1)/lnN,S为总物种数,N为观察到的所有物种的个体总数。(2)多样性指数[19](H)=-∑Pi lnPi,Pi=ni/N,Pi为第i种个体在群落中的比例。
使用SPSS 22.0软件分析不同生境有效夹捕率的差异,采用Kruskal-Wallis H独立样本进行检验,P < 0.05为差异有统计学意义。
2 结果 2.1 鼠类捕获情况及鼠种组成本次调查在11处样地共捕获鼠类102只,经分子鉴定后确定隶属于啮齿目1科5属8种(表 1、2),均属鼠科(Muridae),其中大鼠属(Rattus)3种,白腹鼠属(Niviventer)2种,巨鼠属(Berylmys)、长尾巨鼠属(Leopoldamys)和小鼠属(Mus)各1种。其中华南针毛鼠(N. huang,原中文名拟刺毛鼠)和海南社鼠(N. lotipes)合计占比达73.53%,为该生境小型鼠类优势种;黑缘齿鼠(R. andamanensis)占比为16.67%。区系组成均为东洋界(表 2)。按样地类型区分,农区、景区和林区优势种均为华南针毛鼠,但卡氏小鼠(M. caroli)、黄毛鼠(R. losea)和大足鼠(R. nitidus)仅捕获于农区,小泡巨鼠(L. edwardsi)仅捕获于景区,青毛鼠(B. bowersi)仅捕获于林区。
|

|
>3种不同生境中,景区、农区和林区的平均夹捕率分别为(13.45±3.69)%、(18.24±7.12)%和(7.02±1.25)%,生境间差异无统计学意义(χ2=4.170,P=0.124)。其中农区8号样地夹捕率高达32.43%,林区9号样地夹捕率最低为4.44%。见表 1。
2.2.2 不同鼠种夹捕率在景区、农区华南针毛鼠夹捕率分别为5.59%和7.07%,均为最多鼠种;林区海南社鼠夹捕率最高为3.16%。卡氏小鼠、黄毛鼠、大足鼠均为农区所特有,但夹捕率仅为1.00%左右;青毛鼠和小泡巨鼠夹捕率均 < 0.50%,分别出现在林区和景区。
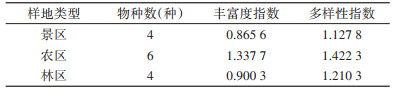
2.3 鼠类生态学指数鼠类丰富度指数以农区样地最高,景区和林区较低,鼠类多样性指数与丰富度指数变化一致。见表 3。
|
通过本次调查,发现广东韶关丹霞山国家自然保护区鼠类分布有1科5属8种。相比于毗邻的广东英德石门台自然保护区[20]、广东杨东山十二度水省级自然保护区[21]和广东连平黄牛石省级自然保护区[22],青毛鼠、小泡巨鼠和黄毛鼠均有分布。而本调查发现的卡氏小鼠、黑缘齿鼠和大足鼠主要在保护区内农区样地有分布,但在杨东山十二度水省级自然保护区的果园、农耕地等区域却未发现以上鼠种[21]。本调查发现华南针毛鼠和海南社鼠为优势种,与其他相近保护区发现的针毛鼠(N. fulvescens)和北社鼠(N. confucianus)可能分别是同物异名,因Ge等[4, 16]使用分子生物学技术对我国白腹鼠属进行系统的重新分类及命名,认为华南区域是华南针毛鼠和海南社鼠的分布区,而不是针毛鼠和北社鼠的分布区。
本调查发现植被类型、生境不同的农区(R样地)、景区(S样地)及林区(F样地)中鼠类的夹捕率、丰富度及多样性指数存在差异,其中农区夹捕率最高,景区次之,林区最低,而物种丰富度及多样性指数林区却高于景区,均低于农区。农区平均夹捕率为18.24%,更有样地高达32.43%,已达到鼠害“大发生”的程度[23],并导致该样地作物近乎绝收。其中不同鼠种夹捕率最高的为华南针毛鼠,而广东省其他市(区),如东莞、江门市调查发现农田中捕获率最高的均为黄毛鼠[17, 23],湛江市多为褐家鼠(R. norvegicus)[24]。华南针毛鼠本应分布于植被茂盛的树林中[10],但本调查中该鼠种捕获率及物种比例均为最高,表明农作物及村民日常食物已将这些野鼠吸引到人类活动范围,造成损失和危害。由此可见,粤北山区农田的主要鼠种结构与其他农区不同,提示需要针对性研究和筛选有效药物和防治方法。
人类活动形成的人为干扰对鼠类群落多样性也具有重要影响,中度干扰假说(intermediate disturbance hypothesis)[25]认为,人类的生活及生产活动在生境中建立了更多的斑块和廊道,增加空间异质性,提高了群落的多样性及种群丰富度。本调查发现,农区(R样地)分布有常年居住村民的村落,土地大多被开垦为农田,人为干扰程度较大、可利用资源较多[26],因此该样地夹捕率、丰富度及多样性指数均最高。观光性旅游活动对一些野生动物的生存造成一定的胁迫,使数量及种类减少[27-28]。本调查景区(S样地)仅节假日客流量较大,人为干扰程度低于农区,游客遗留的垃圾及剩余食物为鼠类提供额外食源,但吸引种类相对单一,造成了景区夹捕率虽高,但丰富度及多样性指数却较农区低的状况。林区(F样地)植物种类较多,面积较大,多为保护区内部未经旅游开发的森林,大多禁止游客或村民入内,人为干扰很小,啮齿类分布相对分散,因此夹捕率较低,但物种丰富度及多样性指数却高于景区,低于农区。此外,农业生产活动会改变区域内的植物覆盖度,造成森林的片段化,对大中型哺乳动物的物种组成造成影响[27-28],而大型动物对人类活动也较为敏感,这可能也导致鼠类天敌动物在农区和景区数量较低,进而鼠类密度较高。
对于尚存在农业生产和景点旅游的自然保护区,鼠类管理需平衡自然生态的需求和生产生活的保障,因而今后需加强定期持续的鼠种类、数量、分布,以及鼠携带寄生虫和病原体的系统监测,并开展针对当地优势害鼠的敏感药物和防治方法的研究,为防控鼠害、保证生产奠定基础。
利益冲突 无
| [1] |
刘铸, 徐艳春, 戎可, 等. 啮齿动物分子系统地理学研究进展[J]. 生态学报, 2014, 34(2): 307-315. Liu Z, Xu YC, Rong K, et al. The current progress in rodents molecular phylogeography[J]. Acta Ecol Sin, 2014, 34(2): 307-315. DOI:10.5846/stxb201304030599 |
| [2] |
魏辅文, 杨奇森, 吴毅, 等. 中国兽类名录(2021版)[J]. 兽类学报, 2021, 41(5): 487-501. Wei FW, Yang QS, Wu Y, et al. Catalogue of mammals in China(2021)[J]. Acta Theriol Sin, 2021, 41(5): 487-501. |
| [3] |
邹发生, 叶冠锋. 广东陆生脊椎动物分布名录[M]. 广州: 广东科技出版社, 2016: 293-296. Zou FS, Ye GF. List of terrestrial vertebrates in Guangdong[M]. Guangzhou: Guangdong Science Technology Press, 2016: 293-296. |
| [4] |
Ge DY, Anderson F, Abramov AV, et al. Molecular phylogeny and morphological diversity of the Niviventer fulvescens species complex with emphasis on species from China[J]. Zool J Linn Soc, 2021, 191(2): 528-547. DOI:10.1093/zoolinnean/zlaa040 |
| [5] |
Cameron GN, Scheel D. Getting warmer: effect of global climate change on distribution of rodents in Texas[J]. J Mammal, 2001, 82(3): 652-680. DOI:10.1644/1545-1542(2001)082<0652:GWEOGC>2.0.CO;2 |
| [6] |
石锐, 李宗智, 高惠, 等. 内蒙古贺兰山啮齿动物群落多样性及其与环境因子关系[J]. 兽类学报, 2019, 39(6): 651-661. Shi R, Li ZZ, Gao H, et al. Rodent diversity and its relationship with environmental factors in Mt. Helan, Inner Mongolia[J]. Acta Theriol Sin, 2019, 39(6): 651-661. DOI:10.16829/j.slxb.150322 |
| [7] |
岳闯, 郭乾伟, 张卓然, 等. 内蒙古典型草原布氏田鼠营养生态位及其种间关系[J]. 兽类学报, 2020, 40(5): 424-434. Yue C, Guo QW, Zhang ZR, et al. Trophic niche of Brandt's voles (Lasiopodomys brandtii) and their interspecific relationships with other common rodents in a typical steppe, Inner Mongolia[J]. Acta Theriol Sin, 2020, 40(5): 424-434. DOI:10.16829/j.slxb.150387 |
| [8] |
朱诚, 马春梅, 张广胜. 中国典型丹霞地貌成因研究[M]. 北京: 科学出版社, 2015: 1-5. Zhu C, Ma CM, Zhang GS. Development mechanisms of typical Danxia landforms in China[M]. Beijing: Science Press, 2015: 1-5. |
| [9] |
Smith AT, Xie Y. A guide to the mammals of China guide to the mammals of China[M]. Changsha: Hunan Education Publishing House, 2009: 160-183.
|
| [10] |
郑智民, 姜志宽, 陈国安. 啮齿动物学[M]. 上海: 上海交通大学出版社, 2008: 165-166. Zheng ZM, Jiang ZK, Chen GA. Rodentology[M]. Shanghai: Shanghai Jiao Tong University Press, 2008: 165-166. |
| [11] |
Robins JH, Hingston M, Matisoo-Smith E, et al. Identifying Rattus species using mitochondrial DNA[J]. Mol Ecol Notes, 2007, 7(5): 717-729. DOI:10.1111/j.1471-8286.2007.01752.x |
| [12] |
Lu L, Chesters D, Zhang W, et al. Small mammal investigation in spotted fever focus with DNA-barcoding and taxonomic implications on rodents species from Hainan of China[J]. PLoS One, 2012, 7(8): e43479. DOI:10.1371/journal.pone.0043479 |
| [13] |
Wei HX, Li FJ, Wang XM, et al. The characterization of complete mitochondrial genome and phylogenetic relationship within Rattus genus (Rodentia: Muridae)[J]. Biochem Syst Ecol, 2017, 71: 179-186. DOI:10.1016/j.bse.2017.01.012 |
| [14] |
Li HT, Kong LM, Wang KY, et al. Molecular phylogeographic analyses and species delimitations reveal that Leopoldamys edwardsi (Rodentia: Muridae) is a species complex[J]. Integr Zool, 2019, 14(5): 494-505. DOI:10.1111/1749-4877.12378 |
| [15] |
Lyu XF, Li HT, Li YY, et al. Characterization of the complete mitochondrial genome of Indochinese forest rat, Rattus andamanensis (Rodentia: Muridae) and its phylogenetic analysis[J]. Mitochondrial DNA B, 2019, 4(1): 1455-1456. DOI:10.1080/23802359.2019.1598811 |
| [16] |
Ge DY, Lu L, Abramov AV, et al. Coalescence models reveal the rise of the white-bellied rat (Niviventer confucianus) following the loss of Asian megafauna[J]. J Mamm Evol, 2019, 26(3): 423-434. DOI:10.1007/s10914-018-9428-y |
| [17] |
姚丹丹, 姜洪雪, 隋晶晶, 等. 广东省江门市捕获鼠类的DNA条形码分析[J]. 中国媒介生物学及控制杂志, 2020, 31(3): 305-309. Yao DD, Jiang HX, Sui JJ, et al. DNA barcoding analysis of rodents captured in Jiangmen, Guangdong province, China[J]. Chin J Vector Biol Control, 2020, 31(3): 305-309. DOI:10.11853/j.issn.1003.8280.2020.03.012 |
| [18] |
Margalef R. Information theory in ecology[J]. Gen Syst, 1958, 3: 36-71. |
| [19] |
Putmann RJ, Wratten SD. Principles of ecology[M]. Worcester: Billing and Sons Limited, 1984: 320-336.
|
| [20] |
吴毅, 易祖盛, 江海声, 等. 广东英德石门台自然保护区啮齿类及翼手类物种多样性研究[J]. 广州大学学报(自然科学版), 2002, 1(2): 21-26. Wu Y, Yi ZS, Jiang HS, et al. Study for species diversity of rodents and bats in Shimentai Nature Reserve, Yingde city, Guangdong[J]. J Guangzhou Univ(Nat Sci Ed), 2002, 1(2): 21-26. DOI:10.3969/j.issn.1671-4229.2002.02.006 |
| [21] |
吴毅, 易祖盛, 江海声. 广东杨东山十二度水省级自然保护区啮齿类、翼手类物种多样性初步研究[J]. 四川动物, 2004, 23(2): 104-107. Wu Y, Yi ZS, Jiang HS. Preliminary study on species biodiversity of rodents and bats in Yangdongshan-Shierdushui Nature Reserve, Guangdong[J]. Sichuan J Zool, 2004, 23(2): 104-107. DOI:10.3969/j.issn.1000-7083.2004.02.006 |
| [22] |
彭红元, 王新财, 江海声, 等. 广东连平黄牛石保护区兽类初步调查[J]. 玉林师范学院学报(自然科学), 2006, 27(3): 109-113. Peng HY, Wang XC, Jiang HS, et al. Preliminary investigation on mannal in Huangniushi NR in Lianping county[J]. J Yulin Teach Coll (Natl Sci), 2006, 27(3): 109-113. DOI:10.3969/j.issn.1004-4671.2006.03.030 |
| [23] |
莫安妮, 温文荣, 姚丹丹, 等. 东莞市农区鼠害的发生动态及综合治理[J]. 热带农业工程, 2014, 38(4): 20-23. Mo AN, Wen WR, Yao DD, et al. Occurrence dynamics and integrated control of farmland rodent in Dongguan[J]. Trop Agric Eng, 2014, 38(4): 20-23. |
| [24] |
姚丹丹, 隋晶晶, 冯志勇. 湛江市褐家鼠种群年龄划分及繁殖特征[J]. 中国媒介生物学及控制杂志, 2016, 27(5): 454-458. Yao DD, Sui JJ, Feng ZY. The population age and reproductive characteristics of Rattus norvegicus in Zhanjiang city[J]. Chin J Vector Biol Control, 2016, 27(5): 454-458. DOI:10.11853/j.issn.1003.8280.2016.05.008 |
| [25] |
Connell JH. Diversity in tropical rain forests and coral reefs: high diversity of trees and corals is maintained only in a nonequilibrium state[J]. Science, 1978, 199(4335): 1302-1310. DOI:10.1126/science.199.4335.1302 |
| [26] |
姚丹丹, 隋晶晶, 刘明津, 等. 农村城镇化对鼠类群落结构的影响研究[J]. 中国媒介生物学及控制杂志, 2014, 25(6): 518-520. Yao DD, Sui JJ, Liu MJ, et al. Study on influence of urbanization on rodent community structure in rural areas[J]. Chin J Vector Biol Control, 2014, 25(6): 518-520. DOI:10.11853/j.issn.1003.4692.2014.06.008 |
| [27] |
张礼标, 郭强, 刘奇, 等. 深圳兽类物种资源调查及其影响因素分析[J]. 兽类学报, 2017, 37(3): 256-265. Zhang LB, Guo Q, Liu Q, et al. Species resources and factors affecting mammals in Shenzhen, Guangdong, China[J]. Acta Theriol Sin, 2017, 37(3): 256-265. DOI:10.16829/j.slxb.201703006 |
| [28] |
Otieno TO, Goheen RJ, Webala PW, et al. Human-and risk-mediated browsing pressure by sympatric antelope in an African savanna[J]. Biol Conserv, 2019, 232: 59-65. DOI:10.1016/j.biocon.2019.01.028 |