 2021, Vol. 32
2021, Vol. 32扩展功能
文章信息
- 刘凯, 姚晓燕, 寸得娇, 张仪, 李兰花
- LIU Kai, YAO Xiao-yan, CUN De-jiao, ZHANG Yi, LI Lan-hua
- 气候环境因子对我国森林革蜱适生区的影响研究
- Effects of climatic and environmental factors on the potential geographic distribution of Dermacentor silvarum in China
- 中国媒介生物学及控制杂志, 2021, 32(6): 732-735
- Chin J Vector Biol & Control, 2021, 32(6): 732-735
- 10.11853/j.issn.1003.8280.2021.06.015
-
文章历史
- 收稿日期: 2021-05-17




2 中国疾病预防控制中心寄生虫病预防控制所, 上海 200025
2 National Institute of Parasitic Diseases, Chinese Center for Disease Control and Prevention, Shanghai 200025, China
蜱是吸血性体外寄生虫,在吸血过程中可传播多种病原体,严重危害人类和动物健康[1]。森林革蜱(Dermacentor silvarum)作为硬蜱中较为常见的一种,可以传播森林脑炎、莱姆病及立克次体病等疾病[2]。本研究拟使用最大熵模型(maximum entropy model,MaxEnt)对森林革蜱在我国的适生区进行分析,并预估未来至2070年气候条件下森林革蜱的分布。
1 材料与方法 1.1 森林革蜱分布点获取在中国知网、万方数据中以“森林革蜱”为检索词进行全文检索,在PubMed、Web of Science数据库中以Dermacentor silvarum〔OR(D. silvarum)AND China〕为检索词进行检索,将所获得的全部中英文文献导入EndNote X9软件查重后逐篇阅读,筛选出包含森林革蜱分布点的文献。从筛选出来的文献中提取森林革蜱地理位置信息;若文献中未给出经纬度信息,则提取调查地点名称信息,通过百度地图(https://map.baidu.com/)中地图开放平台的坐标拾取功能将调查地点转换为经纬度信息。
为避免模型过拟合,将所得的全部分布点坐标导入ArcGIS 10.7软件中,设置半径为5 km的缓冲区,使每个半径为5 km的区域中仅保留1个分布点。
1.2 气候环境因子的获取从全球气候数据网站(https://www.worldclim.org/)中下载当前气候数据(1970—2000年)和2070年气候数据,共包含19个气候因子(bio1~bio19),空间分辨率为5 arc min。地理因子包含坡向、坡度、海拔、植被覆盖率,坡向和坡度可通过ArcGIS 10.7软件处理获得,海拔数据从航天飞机雷达地形测绘任务(shuttle radar topography mission,SRTM)数据(http://srtm.Csi.Cgiar.org/)获得,植被覆盖率由地理空间数据云平台(www.gscloud.cn)获得[3-4]。
将19种气候因子和分布数据导入到ArcGIS 10.7软件中,利用其采样功能提取环境因子,运用SPSS 17.0软件进行相关性检验,若2种环境因子的相关系数 > 0.9,则仅保留生物学意义较大的环境因子纳入后续分析。再通过MaxEnt模型10次运行的平均结果获得单个环境因子变化与物种分布概率关系的响应曲线。
1.3 当前我国森林革蜱分布状况预测将入选的环境气候因子导入到Maxent软件,随机选择70.0%的分布点作为训练集,重复运行10次进行模型构建,用剩余30.0%的分布点作为测试集用于模型验证[5]。
用受试者工作特征曲线(receiver operating characteristic,ROC)下面积(area under curve,AUC)对预测效果进行评价:0.90≤AUC≤1.00为预测效果极好,0.80≤AUC < 0.90为好,0.70≤AUC < 0.80为效果一般,AUC < 0.70为效果较差(失败)[6-8]。最后,用ArcGIS 10.7软件将预测结果可视化,并利用自然划分法将森林革蜱适生区划分为4个等级:分布概率值< 0.12为非适生区,0.12~0.32为低适生区,0.33~0.51为中适生区,> 0.51为高适生区。
1.4 典型路径浓度(RCP)4.5情景下2070年我国森林革蜱分布状况预测从未来温室气体的排放和浓度变化趋势来看,RCP 4.5情景下其在2040年达到峰值,到2080年趋于稳定,这种温室气体排放趋势与中国未来发展趋势一致,符合中国国情[9]。因此本研究拟采用RCP 4.5气候模式预测我国未来气候变化。使用前文中构建的MaxEnt模型,将2070年气候因子、分布数据导入Maxent软件后再次运行,最终得到2070年我国森林革蜱适生分布。
2 结果 2.1 森林革蜱分布点共获取中英文文献839篇,除去重复及数据缺失文献,从109篇中获得218个分布点,经设置半径5 km的缓冲区筛选后,最终保留198个物种分布点用于分析。
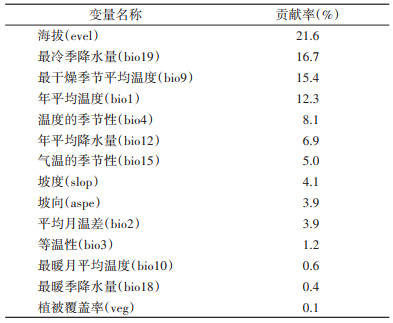
2.2 气候环境因子对森林革蜱分布的影响 2.2.1 气候环境因子筛选结果经相关分析,最终保留海拔、最冷季降水量等14个因子纳入分析,各因子对森林革蜱分布的贡献率见表 1。其中,海拔(evel)、最冷季降水量(bio19)、最干燥季节平均温度(bio9)和年平均温度(bio1)4个因子贡献率超过10.0%。
|
贡献率超过10.0%的因子与森林革蜱分布概率关系的响应曲线见图 1。结果表明,海拔在66.7~3 490.0 m、最冷季降水量在10.2~33.6 mm、最干燥季节平均温度在-27.3~-8.7 ℃、年平均温度在0.4~9.1 ℃时,森林革蜱分布概率较高,分布概率超过高适生区临界值。

|
| 图 1 4种高贡献率气候因子响应曲线 Figure 1 Response curves of four climate factors with high contribution rates |
| |
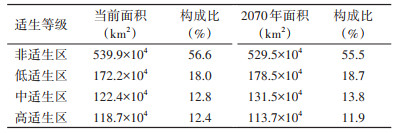
森林革蜱4个等级的适生区范围见图 2,当前和2070年我国森林革蜱适生区面积及所占比例见表 2。结果表明,高适生区主要位于黑龙江、吉林、辽宁、陕西省和新疆维吾尔自治区(新疆)西北部等地区,共118.7×104 km2;中适生区主要分布在内蒙古自治区(内蒙古)东南部、山东和河北省等地区,总共为122.4×104 km2。2070年我国森林革蜱高适生区面积减小5.0×104 km2,损失地区主要位于山西和陕西省;中适生区增加9.1×104 km2,新增地区主要位于山东和陕西省;3类适生区面积总计增加10.4×104 km2。

|
| 图 2 森林革蜱当前适生区及2070年适生区分布变化 Figure 2 The current suitable habitats of Dermacentor silvarum and its distribution changes of suitable habitats in 2070 |
| |
|
本研究表明,我国森林革蜱高适生区主要位于东北、西北等地区,而海拔、最冷季降水量、最干燥季节平均温度、年平均温度是影响森林革蜱分布的主要气候环境因子。赵国平[10]曾应用增强回归树(boosted regression tree,BRT)模型对森林革蜱在全国范围内的分布做出系统调查分析,其研究结果显示森林革蜱的分布范围与本文结果大致相同;气候因子中,最干燥月降水量、最湿季平均温度、气温季节性变化、最干燥季节平均温度、最干燥季节降水量等环境因子对森林革蜱分布影响较大,而海拔、年平均温度等因子对模型的贡献度相对较低,推测出现该差异原因可能为预测软件及原理不同、环境数据来源不同等。另外,本研究侧重于气候变化对森林革蜱分布范围的影响,不考虑湿度等因子,应用了与之不同的模型。我国东北及内蒙古地区最冷季降水量通常在100 mm以下,最干燥季节平均温度通常在-10 ℃以下,如三江平原冬季降水量约为50 mm,冬季平均气温约为-20 ℃,对应森林革蜱高适生区。研究表明,森林革蜱在中国以外地区主要分布于俄罗斯的东南部和蒙古北部[11-13],与我国森林革蜱大部分高适生区相连接。姚文炳和陈国定[14]实验研究表明,温度和湿度对森林革蜱的发育繁殖有重要影响,12~18 ℃、相对湿度55.0%以上时适合其发育繁殖。说明温度和湿度对森林革蜱生长繁殖有较大影响。
预测显示,在RCP 4.5气候模式下,2070年森林革蜱适生区呈扩大趋势。由于全球气候变暖、气温逐年上升、所有地区和所有季节的平均地表温度增加[15],森林革蜱在东北地区的适生区变化不大,但在新疆的适生区大幅减少,山西、陕西、青海省等原为高适生区地带转变为中适生区,原为中适生区的地带转变为低适生区,而在南方的浙江、江西等省非适生区地带逐渐转变为低适生区,其适生区有逐渐向东推移的趋势。上文中提及湿度55.0%以上时有益于森林革蜱生长发育,而我国降水量呈现明显的东部沿海多,西部内陆少,所以降水量可能是导致森林革蜱适生区东移的重要原因。
本研究仍存在如下局限性:首先,从文献中获取的分布点不能代表所有森林革蜱的分布点,未记载有分布的地区可能是无分布或未进行调查;其次,除气候环境因子外,森林革蜱的分布可能受宿主动物分布等因素的影响,由于无法获取相关数据,本研究未分析其他有关因子的影响。因此,未来需进一步开展蜱媒系统监测调查,以更加准确地预测蜱分布范围。
利益冲突 无
| [1] |
刘宇婷, 黄雪玲, 张汉林, 等. 黑龙江省风景区及其周边区域蜱的调查[J]. 现代畜牧科技, 2016(4): 25-26. Liu YT, Huang XL, Zhang HL, et al. Investigation of ticks in Heilongjiang scenic area and its surrounding areas[J]. Mod Anim Husb Sci Technol, 2016(4): 25-26. DOI:10.3969/j.issn.1673-1921.2016.04.016 |
| [2] |
巴音查汗, 岳城, 黄燕, 等. 实验条件下森林革蜱生活史观察[J]. 地方病通报, 2001, 16(2): 74-77. Bayinchahan, Yue C, Huang Y, et al. Observation on the life history of Dermacentor silvarum under the condition of experiment[J]. Endem Dis Bull, 2001, 16(2): 74-77. DOI:10.3969/j.issn.1000-3711.2001.02.029 |
| [3] |
代玉烜, 金瑭, 徐海霞, 等. 基于MaxEnt模型的小黄花茶在中国的适生区域研究[J]. 四川大学学报(自然科学版), 2021, 58(2): 026002. Dai YX, Jin T, Xu HX, et al. Study on the suitable area of Camellia luteoflora Y. K. Li in China based on MaxEnt model[J]. J Sichuan Univ(Nat Sci Ed), 2021, 58(2): 026002. DOI:10.19907/j.0490-6756.2021.026002 |
| [4] |
古丽米拉·克孜尔别克, 邱琴, 海拉提·克孜尔别克. 基于MaxEnt模型的阿勒泰金莲花潜在适生区预测[J]. 江苏农业科学, 2021, 49(4): 82-87. Kezierbieke GLML, Qiu Q, Kezierbieke HLT. Prediction of potential suitable area of Trollius altaicus based on MaxEnt model[J]. Jiangsu Agric Sci, 2021, 49(4): 82-87. DOI:10.15889/j.issn.1002-1302.2021.04.016 |
| [5] |
Escobar LE, Lira-Noriega A, Medina-Vogel G, et al. Potential for spread of the white-nose fungus (Pseudogymnoascus destructans) in the Americas: use of MaxEnt and NicheA to assure strict model transference[J]. Geospat Health, 2014, 9(1): 221-229. DOI:10.4081/gh.2014.19 |
| [6] |
陈智强, 赵增辉, 王远飞, 等. 基于红外相机技术和MaxEnt模型的黑麂(Muntiacus crinifrons)活动节律分析和潜在适生区预测[J]. 生态学报, 2021, 41(9): 3535-3547. Chen ZQ, Zhao ZH, Wang YF, et al. Analysis of activity rhythm and prediction of potential suitable distribution of black muntjac (Muntiacus crinifrons) based on the ITCT and MaxEnt model[J]. Acta Ecol Sin, 2021, 41(9): 3535-3547. DOI:10.5846/stxb201906041185 |
| [7] |
辜云杰, 李晓清, 杨汉波. 基于MaxEnt生态位模型预测桢楠在中国的潜在适宜栽培区[J]. 西北林学院学报, 2021, 36(2): 136-141. Gu YJ, Li XQ, Yang HB. MaxEnt model-based prediction of suitable cultivation area of Phoebe zhennan in China[J]. J Northwest For Univ, 2021, 36(2): 136-141. DOI:10.3969/j.issn.1001-7461.2021.02.20 |
| [8] |
吴瑞婵, 甘淑, 于丽君, 等. 最大熵模型的巴基斯坦遗址预测分布研究[J]. 测绘科学, 2021, 46(3): 96-103. Wu RC, Gan S, Yu LJ, et al. The study of predictive distribution of Pakistani sites based on MaxEnt model[J]. Sci Surv Mapp, 2021, 46(3): 96-103. DOI:10.16251/j.cnki.1009-2307.2021.03.015 |
| [9] |
张蕾, 黄大鹏, 杨冰韵. RCP 4.5情景下中国人口对高温暴露度预估研究[J]. 地理研究, 2016, 35(12): 2238-2248. Zhang L, Huang DP, Yang BY. Future population exposure to high temperature in China under RCP 4.5 scenario[J]. Geogr Res, 2016, 35(12): 2238-2248. DOI:10.11821/dlyj201612004 |
| [10] |
赵国平. 中国蜱类空间分布及其危害预测[D]. 北京: 中国人民解放军军事科学院, 2018. Zhao GP. Spatial distribution of ticks and their hazard prediction in China[D]. Beijing: Academy of Military Science of Chinese People's Liberation Army, 2018. |
| [11] |
Pukhovskaya NM, Morozova OV, Vysochina NP, et al. Prevalence of Borrelia burgdorferi sensu lato and B. miyamotoi in ixodid ticks in the Far East of Russia[J]. Int J Parasitol Parasit Wildl, 2019, 8: 192-202. DOI:10.1016/j.ijppaw.2019.01.005 |
| [12] |
Gaye M, Amanzougaghene N, Laidoudi Y, et al. Hymenopteran parasitoids of hard ticks in western Africa and the Russian far east[J]. Microorganisms, 2020, 8(12): 1992. DOI:10.3390/microorganisms8121992 |
| [13] |
Rubel F, Brugger K, Belova OA, et al. Vectors of disease at the northern distribution limit of the genus Dermacentor in Eurasia: D. reticulatus and D. silvarum[J]. Exp Appl Acarol, 2020, 82(1): 95-123. DOI:10.1007/s10493-020-00533-y |
| [14] |
姚文炳, 陈国定. 森林革蜱生活史与温、湿度的关系[J]. 昆虫学报, 1981, 24(2): 233-236. Yao WB, Chen GD. Duration of the life-history of Dermacentor silvarum olenev. (acari: ixodidae) under different temperatures and humidities[J]. Acta Entomol Sin, 1981, 24(2): 233-236. DOI:10.16380/j.kcxb.1981.02.022 |
| [15] |
Hong CP, Zhang Q, Zhang Y, et al. Impacts of climate change on future air quality and human health in China[J]. Proc Natl Acad Sci USA, 2019, 116(35): 17193-17200. DOI:10.1073/pnas.1812881116 |