 2018, Vol. 53
2018, Vol. 53



2型糖尿病(type 2 diabetes mellitus, T2DM)病因和发病机制目前还不明确, 可能为胰岛β细胞功能缺陷所导致的胰岛素分泌减少(或相对减少), 或胰岛素抵抗所导致的胰岛素在机体内调控葡萄糖代谢能力的下降, 或两者共同存在[1]。在所有糖尿病患者中约占90%。2017年全世界约有4.25亿成年糖尿病患者, 我国18岁及以上人群糖尿病患病率为10.9%, 患者的比例仍在增加[2]。长期高血糖可导致各种大、微血管并发症, 是导致糖尿病患者致残致死的主要因素。因此, 降低2型糖尿病患者血糖新靶点的研究, 对于开发有效安全的降糖药物至关重要。游离脂肪酸不仅是一种产生能量的营养物质, 还可作为细胞外信号分子与多种脂肪酸受体结合。游离脂肪酸受体1 (FFAR1)在胰岛β细胞中大量表达, 具有较强的促进胰岛素分泌的作用, 因此作为潜在的抗糖尿病靶点受到广泛关注。它在促进胰岛素释放、调节葡萄糖稳态、预防及治疗2型糖尿病方面有显著作用。本文就FFAR1在胰岛素分泌调节方面的研究及相关药物开发进展进行综述。
1 FFAR1基因及蛋白结构FFAR1以往又称GPR40, 属于G蛋白偶联受体(GPCRs), 激活后通过一系列信号通路调节细胞功能[3, 4]。FFAR1于1997年被Sawzdargo等[5]发现, 2003年去孤儿化, 是第一个被脱孤的脂肪酸受体[6]。目前已确定的脂肪酸受体有G蛋白偶联受体40 (GPR40)家族, 包括FFAR1 (GPR40)、FFAR2 (GPR43)、FFAR3 (GPR41)及GPR42, 还有其他家族的GPR84、GPR120[7]。GPR40家族的编码基因串联排列于人体19号染色体长臂(19q13.1), 位于CD22基因的下游, 序列具有相似性但各自拥有不同的配基[5]。FFAR1的基因mRNA编码区域长度903 bp, 隶属于长链非编码RNA (lncRNA)类。FFAR1蛋白是具有7次跨膜结构蛋白受体, 含300个氨基酸肽链, 具有高度保守结构。结构上有7个跨膜α螺旋, 且其肽链的C端和连接第5和第6个跨膜螺旋的胞内环上有G蛋白(鸟苷酸结合蛋白)的结合位点。位于跨膜螺旋3和细胞内环2边界处的精氨酸104 (R104)对于FFAR1的正常功能至关重要, R104突变为脯氨酸(R104P)可导致受体功能完全丧失[8]。FFAR1基因中的Arg211His (第211位精、组氨酸)多态性可能导致胰岛素分泌能力的变化[9]。
2 FFAR1表达分布及调控FFAR1在胰岛, 尤其是β细胞中大量表达, 在胰岛α细胞也有表达。FFAR1还表达于各种肠内分泌细胞, 包括分泌胰高血糖素样肽-1 (glucagon-like peptide 1, GLP-1)和多肽YY (PYY)的L细胞[10]、I细胞和K细胞等。另外还表达于脑、胃肠道、肝脏、心脏、骨骼肌、脂肪等组织中。HR2是β细胞特异表达FFAR1的转录增强子, 含有转录因子胰腺十二指肠同源异型盒-1 (pancreas-duodenum homeobox-1, PDX-1)和β细胞E盒转录启动子-2 (β-cell E-box transactivator 2, BETA2)的结合位点, 这两个转录因子与胰岛素基因的转录调控有关。葡萄糖、脂肪酸通过增强PDX-1基因与HR2区域A-box结合, 促进FFAR1基因转录[11]。研究发现, 2型糖尿病患者胰岛的FFAR1表达水平降低[12], 这可能与PDX-1在长期糖脂毒性下活性改变有关[13]。
3 FFAR1在2型糖尿病中的作用及功能FFAR1被中链(6~12个碳原子)和长链(多于12碳原子)脂肪酸激活, 其中包括亚油酸、油酸、棕榈酸等中长链饱和或不饱和游离脂肪酸。FFAR1激活后, 可激活Gq/11, Gs及Gi信号通路, 另外还可激活β-arrestin信号通路[14], 调控胰岛素的分泌调控。胰岛素分泌由许多因素调控, 包括葡萄糖、脂肪酸、肠促胰岛素和交感神经和副交感神经等生理因素[15]。FFAR1对2型糖尿病的调控作用主要表现为参与胰岛素的分泌调节, 包括调控胰岛β的分泌胰岛素功能及细胞增殖; 调控胰岛α细胞分泌胰高血糖素功能; 调控胃肠道内分泌细胞间接促进胰岛素分泌; 控制“脑-脂质传感”参与胰岛素分泌和能量平衡。FFAR1对胰岛素和胰高血糖素都有促分泌作用, 但是在血糖浓度升高时, β细胞生成的胰岛素会抑制胰高血糖素的升血糖作用, 维持血液中葡萄糖稳态。
3.1 FFAR1可调控β细胞分泌胰岛素FFAR1在胰岛β细胞高表达, 2型糖尿病患者的胰岛β细胞内FFAR1表达较低[16]。研究发现FFAR1可放大长链FFAs介导的葡萄糖依赖的胰岛素分泌(glucose stimulated insulin secretion, GSIS)。在过表达胰岛β细胞FFAR1转基因小鼠发现, 胰岛素分泌增加, 葡萄糖耐量增加[17]。Salehi等[18]利用反义寡核苷酸干扰mFFAR1的9~33之间的序列, 抑制FFAR1基因的翻译, 诱导产生基因沉默FFAR1-/-小鼠, 表现出亚油酸(linoleic acid, LA)诱导的葡萄糖依赖的胰岛素释放减弱。Sabrautzki等[19]用胰岛FFAR1基因的R258W错义点突变小鼠, 给予棕榈酸酯和FFAR1激动剂, 发现FFAR1 (R258W)突变小鼠失去FFAs促进胰岛素释放作用, 但高脂诱导的葡萄糖耐受不良得到改善。20-羟-二十烷四烯酸(20-HETE)可激活FFAR1并形成一个正反馈环, 增强葡萄糖刺激的胰岛素分泌[20]。FFAR1信号可能通过增强线粒体呼吸来影响棕榈酸刺激的胰岛素分泌[21]。
胰岛素在胰岛β细胞中合成储存并释放, 当血液中葡萄糖含量升高时, β细胞膜上的葡萄糖转运体2 (glucose transporter-2, GLUT-2)把血液中高浓度葡萄糖转运到β细胞内, 在葡糖激酶(glucokinase, GK)的作用下, 葡萄糖被磷酸化为葡糖-6-磷酸(glucose- 6-phosphate, G-6-P)。GK只有在血糖浓度大于5 mmol·L-1时发挥作用[22], 使胰岛素只在需要时分泌从而降低血糖。G-6-P继续水解为丙酮酸, 丙酮酸进入线粒体氧化脱羧生成乙酰-CoA, 然后进入三羧酸循环(TCA), 消耗ADP生成ATP, 即ATP/ADP升高, 细胞膜上的ATP敏感的K+ (KATP)通道关闭, 膜去极化, 电压门控L型Ca2+ (L-type Ca2+-channel, LTCC)通道打开, Ca2+内流, 细胞内Ca2+浓度([Ca2+]i)升高, 从而使胰岛素经β细胞分泌颗粒释放到胞外。电压依赖的K+ (Kv)通道开放, K+外流, β细胞膜复极化, LTCC通道关闭, 保持动作电位短暂, 限制Ca2+内流及胰岛素释放的时间长度。
FFAR1促进胰岛素分泌的信号机制(图 1)为, FFAR1激活后可同时经G蛋白及β-arrestin信号通路传导, 内源性及外源性配体可经不同通路发挥作用[14]。FFAR1通过Gαq/11亚基磷酸化激活磷脂酶C (phospholipase C, PLC)[23], 催化4, 5-二磷酸磷脂酰肌醇(phosphatidylinositol-4, 5-bisphosphate, PIP2)产生第二信使1, 4, 5-三磷酸肌醇(inositol-1, 4, 5-triphosphate, IP3)和甘油二酯(diacyl glycerol, DAG)。IP3激活内质网上的IP3受体(IP3R), Ca2+由内质网释放, DAG可激活蛋白激酶C (protein kinase C, PKC), 或直接促进分泌颗粒与膜融合, 或激活蛋白激酶D1 (protein kinase D1, PKD1), 使F-肌动蛋白解聚, 帮助分泌颗粒在胞内转移。另外, FFAR1也可通过Gαs亚基激活腺苷酸环化酶(adenylyl cyclase, AC), 产生环磷酸腺苷(cyclic adenosine monophosphate, cAMP), 升高细胞内cAMP水平, 拮抗电压门控钾离子通道的活性, 膜去极化时间延长, 增强Ca2+由LTCC内流, 维持细胞内的高水平Ca2+, 促进胰岛素释放[24]。放大途径也是在葡萄糖达到一定浓度(大于1 mmol·L-1[25])条件下进行的, 可避免胰岛素过度分泌引起高胰岛素血症和低血糖症。FFAR1信号也可通过过氧化物酶体增殖物激活受体PPARγ来介导, 二者可以作为一种整合的双受体信号转导途径发挥抗糖尿病作用[26]。Mancini等[14]研究发现FFAR1还可通过招募β-arrestin 2通路, 调节胰岛素释放。不同配体引发受体不同的构象改变, 有些构象利于G蛋白结合, 有些构象则利于β-arrestin结合, 从而分别激活配体特异的下游信号传递信号。内源性配体棕榈酸和油酸倾向于激活Gαq信号, 而合成的FFAR1激动剂TAK-875主要促进β-arrestin 2介导的信号传导, 具有受体偏向性。

|
Figure 1 FFAR1 signaling in pancreatic β-cells. FFAs: Free fatty acids; FFAR1: Free fatty acid receptor 1; PLC: Phospholipase C; PIP2: Phosphatidylinositol-4, 5-bisphosphate; IP3: Inositol-1, 4, 5-triphosphate; IP3R: IP3 receptor; ER: Endoplasmic reticulum; DAG: Diacyl glycerol; PKC: Protein kinase C; PKD1: Protein kinase D1; AC: Adenylyl cyclase; cAMP: Cyclic adenosine monophosphate; PKA: Protein kinase A; [Ca2+]i: Intracellular Ca2+ concentration |
研究还发现FFAR1在介导脂肪酸导致的胰岛β细胞胰岛素分泌中具有双向性, 短期急性给予可促进胰岛素分泌, 而长期作用则发挥相反作用[27]。FFAs对GSIS的作用是双向的, 短暂大量的FFAs可增强GSIS, 长时间高浓度FFAs则会产生脂毒性。FFAR1至少部分介导短期FFAs对GSIS的促进作用, 而FFAR1是否也介导长期FFAs对GSIS的抑制作用则存在争议。一方面, Steneberg等[28]发现FFAR1转基因高表达小鼠在正常饮食条件下会发生葡萄糖耐受和高胰岛素血症, 而FFAR1基因敲除小鼠在高脂饮食条件下, 没有发生胰岛素抵抗, 认为FFAR1同时介导短期FFAs的促胰岛素和长期FFAs的抑制胰岛素作用。另一方面, Wu等[29]体外实验发现, 长期FFAs刺激对FFAR1基因沉默的小鼠胰岛素瘤细胞系β-TC6有显著抑制作用。Lan等[30]体内实验表明, 野生小鼠和FFAR1基因完全敲除小鼠在高脂喂养下具有相似的高胰岛素血症, 葡萄糖耐量和胰岛素耐量无差异, 认为慢性脂肪酸引起的促胰岛β细胞凋亡作用与FFAR1无关。Wang等[31]认为过量脂肪酸对胰岛β细胞的损伤与线粒体外膜破裂有关。
3.2 FFAR1可调控α细胞生成胰高血糖素胰高血糖素在血糖浓度低时, 由α细胞分泌, 促进肝脏糖原转变为葡萄糖, 抑制糖原生成。Flodgren等[32]发现, 在中低葡萄糖浓度下, FFAR1具有促进胰高血糖素生成, 升高血糖的作用。研究表明, 对小鼠胰岛进行复染, 用共聚焦显微镜观察FFAR1和胰高血糖素分布, 发现除了胰岛β细胞, 在分泌胰高血糖素的α细胞中也可检测到FFAR1表达。仓鼠胰高血糖素瘤细胞株In-R1-G9也可内源性表达FFAR1, LA刺激过表达FFAR1的细胞, 出现磷脂酰肌醇(PI)水解和胰高血糖素分泌剂量依赖性增加; 在1及8.3 mmol·L-1葡萄糖条件下, LA处理分离的胰岛组织, 也可见胰高血糖素分泌增加。反之, LA刺激反义寡核苷酸沉默FFAR1表达的细胞, 胰高血糖素分泌减少, 表明FFAR1在α细胞表达并发挥促胰高血糖素分泌作用。Verma等[33]发现, 在高葡萄糖浓度下, 大鼠胰岛分泌胰高血糖素降低, 激动FFAR1后更加抑制了胰高血糖素分泌。
3.3 FFAR1通过调控胃肠道内分泌细胞间接促进胰岛素分泌胃肠道内分泌细胞中有大量FFAR1存在, 并可促进肠促胰岛素分泌。肠促胰岛素是一组具有降血糖作用胃肠道激素的统称, 在糖代谢调节中起重要作用。Edfalk等[10]用lacZ报告基因插入小鼠FFAR1基因, 结果在胃幽门部、十二指肠、结肠、空肠、回肠细胞均观察到明显的β-半乳糖苷酶显色反应, 进而发现FFAR1在胃肠道内分泌细胞表达, 包括分泌GLP-1的L细胞、分泌GIP的K细胞、分泌胆囊收缩素(CCK)的肠内分泌细胞I细胞。GLP-1及GIP是肠促胰岛素主要成员, 具有促进胰岛素分泌、减少肝脏糖异生、抑制胰高血糖素释放、延缓胃排空、抑制食欲的作用。研究发现, FFAR1可介导FFAs诱导的GLP-1[34-36]及GIP分泌, 使胰岛素分泌增加[10]。高脂饲养的FFAR1基因敲除小鼠在FFAs刺激下, 与正常小鼠比较可见胰岛素水平下降, 同时伴有GLP-1及GIP表达减少[32]。在胃幽门、十二指肠也存在调节FFAR1表达的Ipf1/Pdx1基因[10]。表明FFAR1直接作用于β细胞促进胰岛素分泌之外, 还可调节GLP-1及GIP的肠促胰岛素作用, 间接介导FFAs促进的胰岛素分泌[10]。
肠内分泌细胞I细胞分泌的CCK也可以调节胰岛素分泌[37]。Liou等[38]用eGFP与CCK基因融合, 标记表达CCK的I细胞。结果显示, 在表达CCK的细胞中也检测到FFAR1的表达, 表达量远大于不表达CCK的细胞。长链FFAs可增加FFAR1过表达I细胞的CCK分泌及[Ca2+]i, 而在无FFAR1表达的I细胞CCK分泌减少, [Ca2+]i降低。这表明FFAR1还可诱导CCK的分泌, 间接促进胰岛素的产生[38]。
3.4 FFAR1可能通过脑调节胰岛素分泌FFAR1还广泛分布在人脑的各个区域, 目前研究发现可能与胰岛素分泌及糖脂代谢调节有关。一般认为FFAs不是神经代谢的初级燃料, 故其在脑代谢中的作用一直不被认可。但越来越多的证据表明, FFAs可在中枢神经系统作为细胞信使, 向脂肪酸敏感神经元传递身体的能量状况信号, 即“脑-脂质传感”[39], 从而对饮食行为、肝糖生成、胰岛素分泌产生影响。据报道, FFAR1在人脑中各个区域(包括下丘脑)也普遍存在[40]。下丘脑神经元和胰岛β细胞的共同点是都能对FFAs产生应答, 并与能量平衡的调节有关[41]。“脑-脂质传感”可能参与胰岛素分泌和能量平衡的控制。胰岛中副交感神经和交感神经的分布广泛, 提示下丘脑和胰腺的亲密关系。副交感神经末梢释放乙酰胆碱, 激活胰岛β细胞表面M2受体, 促进胰岛素通过甘油二酯-蛋白激酶C (DAG-PKC)依赖途径释放。交感神经激动β-肾上腺素能受体和α2-肾上腺素能受体, β-受体活性增加增强胰岛素释放, 而α2-受体活性增加减少胰岛素释放。两种通路通过cAMP作用, 分别导致cAMP水平的减少或增多[15]。但FFAR1在脑组织表达与胰岛素分泌的具体关联及作用尚不明确, 对FFAR1作用的深入理解可能为代谢性疾病的防治提供重要线索。
3.5 FFAR1与其他组织FFAR1还表达于肝脏、心脏、骨骼肌、脂肪等组织中。肝脏、脂肪、骨骼肌组织在全身葡萄糖稳态方面的作用极其重要, 它是血糖消耗、转化和储存的场所, 也是调控刺激胰岛素分泌的主要器官。
研究发现, FFAs通过抑制胰岛素信号通路和葡萄糖转运导致骨骼肌胰岛素抵抗[42]。FFAR1在骨骼肌中也有表达。短期FFAs升高会损害基础状态骨骼肌的葡萄糖利用[5], 长期高水平FFAs或高水平胰岛素, 会促进骨骼肌糖摄取, 增加骨骼肌内脂肪和糖原的积累, 导致骨骼肌胰岛素抵抗及2型糖尿病的发生[43]。
4 以FFAR1为靶点的药物开发FFAR1在促进胰岛素分泌方面的作用引起广泛关注, 具有很大的药物开发潜力。目前针对该靶点的激动剂包括完全激动剂、选择性激动剂、变构激动剂。当前的临床证据表明, FFAR1激动剂能够通过激活FFAR1, 调节胰岛素分泌, 并具有葡萄糖依赖性, 即血糖上升才能促进胰岛素释放, 降低餐后血糖而且没有低血糖的风险, 因此优于磺酰脲类和格列奈类药物; FFAR1激动剂能够调节GLP-1释放, 在促进胰岛素分泌的同时能降低体重及保护β细胞, 没有增重的风险, 因此较噻唑烷二酮类和PPAR激动剂药物有优势。但截止到2017年底, 相关FFAR1的药物开发均仅停留在临床研究阶段, 尚未有药物上市。
日本武田制药(Takeda)的FFAR1激动剂Fasiglifam (也称TAK-875)是第一个进入Ⅲ期临床试验的药物, 同时也是FFAR1的变构激活剂[44], 2013年12月因药物肝毒性而停止研发。有研究发现其诱导肝脏损害与其诱发的氧化应激导致的活性氧生成增加有关[45]。美国礼来(Eli Lilly)公司的LY 2922470是一种FFAR1 Gαq亚基选择性激动剂[46], 目前已完成临床概念性验证研究与临床Ⅰ期试验, 包括单剂量给药及患者重复剂量给药试验。
FFAR1的选择性激动剂LY3104607、变构激活剂Fasiglifam/TAK-875、部分激动剂MK-8666[47]、完全变构激动剂AgoPAMs[48]与FFAR1结合的晶体结构也得到解析。FFAR1激动剂也有一些结构优化和修饰改造产物, 具有较好的效果[49, 50], 还有一些天然产物也具有FFAR1激动作用[51, 52]。目前在临床阶段的主要FFAR1激动剂见表 1[46, 53-56]。
| Table 1 The main FFAR1 agonist being studied in clinical research |

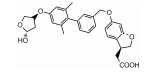

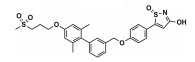
目前, 基础和临床研究均提示FFAR1是抗糖尿病治疗的潜在靶点, 但有关FFAR1的研究仍存在争议, 到目前为止尚无药物上市。一方面, FFAR1是否介导长期高浓度FFAs导致的β细胞脂毒性, 尚不明确; 另一方面, 激动FFAR1不仅可促进胰岛素分泌, 还可促进胰高血糖素分泌, 而这两种激素作用相反。虽然目前还没有定论, 但FFAR1激动剂在临床试验上的良好结果, 为选择FFAR1激动剂而不是拮抗剂用于抗糖尿病治疗提供了有力依据。目前, FFAR1的选择性激动剂、完全激动剂、拮抗剂的研究都已取得一定进展, 但药效和安全性仍存在问题。FFAR1作为2型糖尿病治疗的靶点, 其药理作用及机制仍需要深入研究。
| [1] | Chinese Diabetes Society. Guidelines for prevention and treatment of type 2 diabetes in China (2017 version)[J]. Chin J Diebetes Mellitus (中华糖尿病杂志), 2018, 10: 4–67. |
| [2] | Wang L, Gao P, Zhang M, et al. Prevalence and ethnic pattern of diabetes and prediabetes in China in 2013[J]. JAMA, 2017, 317: 2515–2523. DOI:10.1001/jama.2017.7596 |
| [3] | Aizawa F, Nishinaka T, Yamashita T, et al. GPR40/FFAR1 deficient mice increase noradrenaline levels in the brain and exhibit abnormal behavior[J]. J Pharmacol Sci, 2016, 132: 249–254. DOI:10.1016/j.jphs.2016.09.007 |
| [4] | Stein DT, Esser V, Stevenson BE, et al. Essentiality of circulating fatty acids for glucose-stimulated insulin secretion in the fasted rat[J]. J Clin Invest, 1996, 97: 2728–2735. DOI:10.1172/JCI118727 |
| [5] | Sawzdargo M, George SR, Nguyen TV, et al. A cluster of four novel human G protein-coupled receptor genes occurring in close proximity to CD22 gene on chromosome 19q13.1[J]. Biochem Biophys Res Commun, 1997, 239: 543–547. DOI:10.1006/bbrc.1997.7513 |
| [6] | Briscoe CP, Tadayyon M, Andrews EJ, et al. The orphan G protein-coupled receptor GPR40 is activated by medium and long-chain fatty acids[J]. J Biol Chem, 2003, 278: 11303–11311. DOI:10.1074/jbc.M211495200 |
| [7] | Liaw CW, Connolly DT. Sequence polymorphisms provide a common consensus sequence for GPR41 and GPR42[J]. DNA Cell Biol, 2009, 28: 555–560. DOI:10.1089/dna.2009.0916 |
| [8] | Guo S, Zhang J, Zhang S, et al. A single amino acid mutation (R104P) in the E/DRY motif of GPR40 impairs receptor function[J]. PLoS One, 2015, 10: e0141303. DOI:10.1371/journal.pone.0141303 |
| [9] | Ogawa T, Hirose H, Miyashita K, et al. GPR40 gene Arg211His polymorphism may contribute to the variation of insulin secretory capacity in Japanese men[J]. Metabolism, 2005, 54: 296–299. DOI:10.1016/j.metabol.2004.09.008 |
| [10] | Edfalk S, Steneberg P, Edlund H, et al. Gpr40 is expressed in enteroendocrine cells and mediates free fatty acid stimulation of incretin secretion[J]. Diabetes, 2008, 57: 2280–2287. DOI:10.2337/db08-0307 |
| [11] | Bartoovshifman R, Ridner G, Bahar K, et al. Regulation of the gene encoding GPR40, a fatty acid receptor expressed selectively in pancreatic β cells[J]. J Biol Chem, 2007, 282: 23561–23571. DOI:10.1074/jbc.M702115200 |
| [12] | Guerra SD, Bugliani M, Daleo V, et al. G-protein-coupled receptor 40 (GPR40) expression and its regulation in human pancreatic islets: the role of type 2 diabetes and fatty acids[J]. Nutr Metab Cardiovasc Dis, 2010, 20: 22–25. DOI:10.1016/j.numecd.2009.02.008 |
| [13] | Poitout V, Amyot J, Semache M, et al. Glucolipotoxicity of the pancreatic beta cell[J]. Biochim Biophys Acta, 2010, 1801: 289–298. DOI:10.1016/j.bbalip.2009.08.006 |
| [14] | Mancini A, Bertrand G, Vivot K, et al. β-Arrestin recruitment and biased agonism at free fatty acid receptor 1[J]. J Biol Chem, 2015, 290: 21131–21140. DOI:10.1074/jbc.M115.644450 |
| [15] | Mizgier ML, Casas M, Contrerasferrat A, et al. Potential role of skeletal muscle glucose metabolism on the regulation of insulin secretion[J]. Obes Rev, 2014, 15: 587–597. DOI:10.1111/obr.2014.15.issue-7 |
| [16] | Tomita T, Masuzaki H, Iwakura H, et al. Expression of the gene for a membrane-bound fatty acid receptor in the pancreas and islet cell tumours in humans: evidence for GPR40 expression in pancreatic beta cells and implications for insulin secretion[J]. Diabetologia, 2006, 49: 962–968. DOI:10.1007/s00125-006-0193-8 |
| [17] | Nagasumi K, Esaki R, Iwachidow K, et al. Overexpression of GPR40 in pancreatic β-cells augments glucose-stimulated insulin secretion and improves glucose tolerance in normal and diabetic mice[J]. Diabetes, 2009, 58: 1067–1076. DOI:10.2337/db08-1233 |
| [18] | Salehi SA, Flodgren E, Nilsson NE, et al. Free fatty acid receptor 1 (FFA1R/GPR40) and its involvement in fatty-acid- stimulated insulin secretion[J]. Cell Tissue Res, 2005, 322: 207–215. DOI:10.1007/s00441-005-0017-z |
| [19] | Sabrautzki S, Kaiser G, Przemeck GK, et al. Point mutation of FFAR1 abrogates fatty acid-dependent insulin secretion, but protects against HFD-induced glucose intolerance[J]. Mol Metab, 2017, 6: 1304–1312. DOI:10.1016/j.molmet.2017.07.007 |
| [20] | Tunaru S, Bonnavion R, Brandenburger I, et al. 20-HETE promotes glucose-stimulated insulin secretion in an autocrine manner through FFAR1[J]. Nat Commun, 2018, 9: 177. DOI:10.1038/s41467-017-02539-4 |
| [21] | Kristinsson H, Bergsten P, Sargsyan E, et al. Free fatty acid receptor 1 (FFAR1/GPR40) signaling affects insulin secretion by enhancing mitochondrial respiration during palmitate exposure[J]. Biochim Biophys Acta, 2015, 1853: 3248–3257. DOI:10.1016/j.bbamcr.2015.09.022 |
| [22] | Matschinsky FM, Glaser B, Magnuson MA, et al. Pancreatic beta-cell glucokinase: closing the gap between theoretical concepts and experimental realities[J]. Diabetes, 1998, 47: 307–315. DOI:10.2337/diabetes.47.3.307 |
| [23] | Yamada H, Yoshida M, Ito K, et al. Potentiation of glucose- stimulated insulin secretion by the GPR40-PLC-TRPC pathway in pancreatic β-cells[J]. Sci Rep, 2016, 6: 25912. DOI:10.1038/srep25912 |
| [24] | Offermanns S. G-proteins as transducers in transmembrane signalling[J]. Prog Biophys Mol Biol, 2003, 83: 101–130. DOI:10.1016/S0079-6107(03)00052-X |
| [25] | Henquin J. Triggering and amplifying pathways of regulation of insulin secretion by glucose[J]. Diabetes, 2000, 49: 1751–1760. DOI:10.2337/diabetes.49.11.1751 |
| [26] | Wang S, Awad KS, Elinoff JM, et al. G protein-coupled receptor 40 (GPR40) and peroxisome proliferator-activated receptor γ (PPARγ): an integrated two-receptor signaling pathway[J]. J Biol Chem, 2015, 290: 19544–19557. DOI:10.1074/jbc.M115.638924 |
| [27] | Kristinsson H, Smith DM, Bergsten P, et al. FFAR1 is involved in both the acute and chronic effects of palmitate on insulin secretion[J]. Endocrinology, 2013, 154: 4078–4088. DOI:10.1210/en.2013-1352 |
| [28] | Steneberg P, Rubins N, Bartoovshifman R, et al. The FFA receptor GPR40 links hyperinsulinemia, hepatic steatosis, and impaired glucose homeostasis in mouse[J]. Cell Metab, 2005, 1: 245–258. DOI:10.1016/j.cmet.2005.03.007 |
| [29] | Wu P, Yang L, Shen X, et al. The relationship between GPR40 and lipotoxicity of the pancreatic β-cells as well as the effect of pioglitazone[J]. Biochem Biophys Res Commun, 2010, 403: 36–39. DOI:10.1016/j.bbrc.2010.10.105 |
| [30] | Lan H, Hoos LM, Liu L, et al. Lack of FFAR1/GPR40 does not protect mice from high-fat diet-induced metabolic disease[J]. Diabetes, 2008, 57: 2999–3006. DOI:10.2337/db08-0596 |
| [31] | Wang Y, Liu SN, Shen ZF. Dynamic associations of mitochondria-endoplasmic reticulum in maintenance of pancreatic beta cell homeostasis[J]. Acta Pharm Sin (药学学报), 2017, 52: 667–672. |
| [32] | Flodgren E, Olde B, Meiduteabaraviciene S, et al. GPR40 is expressed in glucagon producing cells and affects glucagon secretion[J]. Biochem Biophys Res Commun, 2007, 354: 240–245. DOI:10.1016/j.bbrc.2006.12.193 |
| [33] | Verma MK, Biswas S, Chandravanshi B, et al. A novel GPR40 agonist, CNX-011-67, suppresses glucagon secretion in pancreatic islets under chronic glucolipotoxic conditions in vitro[J]. BMC Res Notes, 2014, 7: 595. DOI:10.1186/1756-0500-7-595 |
| [34] | Hauge M, Vestmar MA, Husted AS, et al. GPR40 (FFAR1)- combined Gs and Gq signaling in vitro is associated with robust incretin secretagogue action ex vivo and in vivo[J]. Mol Metab, 2015, 4: 3–14. DOI:10.1016/j.molmet.2014.10.002 |
| [35] | Guo D, Li D, Ning M, et al. Yhhu4488, a novel GPR40 agonist, promotes GLP-1 secretion and exerts anti-diabetic effect in rodent models[J]. Biochem Biophys Res Commun, 2015, 466: 740–747. DOI:10.1016/j.bbrc.2015.09.130 |
| [36] | Li A, Wang Q, Dinh TT, et al. Mercaptoacetate blocks fatty acid-induced GLP-1 secretion in male rats by directly antagonizing GPR40 fatty acid receptors[J]. Am J Physiol Regul Integr Comp Physiol, 2016, 310: R724–R732. DOI:10.1152/ajpregu.00387.2015 |
| [37] | Nauck MA, Homberger E, Siegel EG, et al. Incretin effects of increasing glucose loads in man calculated from venous insulin and C-peptide responses[J]. J Clin Endocrinol Metab, 1986, 63: 492–498. DOI:10.1210/jcem-63-2-492 |
| [38] | Liou AP, Lu X, Sei Y, et al. The G-protein-coupled receptor GPR40 directly mediates long-chain fatty acid-induced secretion of cholecystokinin[J]. Gastroenterology, 2011, 140: 903–912. DOI:10.1053/j.gastro.2010.10.012 |
| [39] | Moulle VS, Picard A, Foll CL, et al. Lipid sensing in the brain and regulation of energy balance[J]. Diabetes Metab, 2014, 40: 29–33. DOI:10.1016/j.diabet.2013.10.001 |
| [40] | Yamashima T. Dual effects of the non-esterified fatty acid receptor 'GPR40' for human health[J]. Prog Lipid Res, 2015, 58: 40–50. DOI:10.1016/j.plipres.2015.01.002 |
| [41] | Dragano NR, Solon C, Ramalho AF, et al. Polyunsaturated fatty acid receptors, GPR40 and GPR120, are expressed in the hypothalamus and control energy homeostasis and inflammation[J]. J Neuroinflammation, 2017, 14: 91. DOI:10.1186/s12974-017-0869-7 |
| [42] | Griffin ME, Marcucci M, Cline GW, et al. Free fatty acid- induced insulin resistance is associated with activation of protein kinase C theta and alterations in the insulin signaling cascade[J]. Diabetes, 1999, 48: 1270–1274. DOI:10.2337/diabetes.48.6.1270 |
| [43] | Jenkins AB, Storlien LH, Chisholm DJ, et al. Effects of nonesterified fatty acid availability on tissue-specific glucose utilization in rats in vivo[J]. J Clin Invest, 1988, 82: 293–299. DOI:10.1172/JCI113586 |
| [44] | Kaku K, Enya K, Nakaya R, et al. Long-term safety and efficacy of fasiglifam (TAK-875), a G-protein-coupled receptor 40 agonist, as monotherapy and combination therapy in Japanese patients with type 2 diabetes: a 52-week open-label phase Ⅲ study[J]. Diabetes Obes Metab, 2016, 18: 925–929. DOI:10.1111/dom.12693 |
| [45] | Kim M, Gu GJ, Koh YS, et al. Fasiglifam (TAK-875), a G protein-coupled receptor 40 (GPR40) agonist, may induce hepatotoxicity through reactive oxygen species generation in a GPR40-dependent manner[J]. Biomol Ther, 2018. DOI:10.4062/biomolther.2017.225 |
| [46] | Hamdouchi C, Kahl SD, Lewis AP, et al. The discovery, preclinical, and early clinical development of potent and selective GPR40 agonists for the treatment of type 2 diabetes mellitus (LY2881835, LY2922083, and LY2922470)[J]. J Med Chem, 2016, 59: 10891–10916. DOI:10.1021/acs.jmedchem.6b00892 |
| [47] | Krug AW, Vaddady P, Railkar RA, et al. Leveraging a clinical phase Ib proof-of-concept study for the GPR40 agonist MK-8666 in patients with type 2 diabetes for model-informed Phase Ⅱ dose selection[J]. Clin Transl Sci, 2017, 10: 404–411. DOI:10.1111/cts.2017.10.issue-5 |
| [48] | Pachanski M, Kirkland M, Kosinski DT, et al. GPR40 partial agonists and AgoPAMs: differentiating effects on glucose and hormonal secretions in the rodent[J]. PLoS One, 2017, 12: e0186033. DOI:10.1371/journal.pone.0186033 |
| [49] | Krasavin M, Lukin A, Bagnyukova D, et al. Polar aromatic periphery increases agonist potency of spirocyclic free fatty acid receptor (GPR40) agonists inspired by LY2881835[J]. Eur J Med Chem, 2017, 127: 357–368. DOI:10.1016/j.ejmech.2017.01.005 |
| [50] | Badolato M, Carullo G, Perri M, et al. Quercetin/oleic acid- based G-protein-coupled receptor 40 ligands as new insulin secretion modulators[J]. Future Med Chem, 2017, 9: 1873–1885. DOI:10.4155/fmc-2017-0113 |
| [51] | Feng XT, Duan HM, Li SL. Protective role of Pollen Typhae total flavone against the palmitic acid-induced impairment of glucose-stimulated insulin secretion involving GPR40 signaling in INS-1 cells[J]. Int J Mol Med, 2017, 40: 922–930. DOI:10.3892/ijmm.2017.3070 |
| [52] | Krasavin M, Lukin A, Bakholdina A, et al. Continued SAR exploration of 1, 2, 4-thiadiazole-containing scaffolds in the design of free fatty acid receptor 1 (GPR40) agonists[J]. Eur J Med Chem, 2017, 140: 229–238. DOI:10.1016/j.ejmech.2017.09.019 |
| [53] | Kim S, Kim DH, Kim Y, et al. HD047703, a new promising anti-diabetic drug candidate: in vivo preclinical studies[J]. Biomol Ther, 2014, 22: 400–405. DOI:10.4062/biomolther.2014.035 |
| [54] | Li Z, Xu X, Huang W, et al. Free fatty acid receptor 1 (FFAR1) as an emerging therapeutic target for type 2 diabetes mellitus: recent progress and prevailing challenges[J]. Med Res Rev, 2018, 38: 381–425. DOI:10.1002/med.21441 |
| [55] | Miller C, Pachanski MJ, Kirkland ME, et al. GPR40 partial agonist MK-2305 lower fasting glucose in the Goto Kakizaki rat via suppression of endogenous glucose production[J]. PLoS One, 2017, 12: e0176182. DOI:10.1371/journal.pone.0176182 |
| [56] | Tsuda N, Kawaji A, Sato T, et al. A novel free fatty acid receptor 1 (GPR40/FFAR1) agonist, MR1704, enhances glucose- dependent insulin secretion and improves glucose homeostasis in rats[J]. Pharmacol Res Perspect, 2017. DOI:10.1002/prp2.340 |