 2018, Vol. 53
2018, Vol. 53



高脂血症是诱发心血管疾病的主要危险因素之一, 控制血脂的升高对于预防心血管疾病具有十分重要的意义[1, 2]。临床上高脂血症是由血清中甘油三酯(TG)、总胆固醇(TC)和低密度脂蛋白胆固醇(LDL-C)水平升高以及高密度脂蛋白胆固醇(HDL-C)水平降低来判断[3, 4]。尽管许多脂代谢紊乱患者通过调节饮食和运动来控制血脂, 但收效甚微, 最终还是依靠长期服用降脂药。越来越多的降脂药被开发和应用于治疗脂代谢紊乱。他汀类、贝特类、烟酸类、胆固醇吸收抑制剂是国际上主流的降脂药。目前所用的降脂药无一例外均有多种不良反应, 比如肝功能异常、恶心、胆囊炎、肌炎等[5]。近年来, 许多研究表明高脂饮食与肠道菌群的改变密切相关, 肠道菌群中的肠杆菌、肠球菌、双歧杆菌、乳酸杆酸等多种菌属与脂代谢有着密切的关系, 而脂代谢异常会直接影响肠道菌群的数量和分布。目前无论是体外实验还是动物实验均证实, 脂代谢紊乱可导致肠道菌群失调, 而肠道菌群失调又可进一步加重脂代谢紊乱。肠道菌群与脂代谢紊乱的关系已成为近年来研究的热点。
近来, 迅速发展的代谢组学技术为探索肠道菌群如何影响宿主生理和如何利用肠道菌群开发新的治疗和营养策略提供新视角。代谢组学技术能为微生物和宿主的相互作用提供一个快照, 为肠道菌群对人类健康与疾病的影响提供分子基础[6]。例如, 一系列关于人、大鼠、无菌小鼠的研究为评估肠道菌群结构和功能提供了代谢轮廓信息[7-10]。这些结果为肠道菌群在代谢类疾病发展中的作用提供了重要信息, 这可能对脂代谢紊乱疾病的预防和治疗策略起到重要作用。以上研究表明结合代谢组学技术研究肠道菌群在脂代谢紊乱发生发展过程中的作用是非常有必要的。
在目前的研究中, 高脂血症是由高脂肪饮食诱导, 其中高脂饮食诱导的金黄地鼠高脂血症模型与人更为相似, 是研究人员了解脂代谢紊乱特征、探索其发生发展机制的最佳小动物模型[11]。但关于金黄地鼠肠道内容物和粪便的研究鲜有报道, 这是对研发降血脂药物的重要动物模型的一个缺失。综上, 本研究采用了基于1H NMR的代谢组学技术和多元统计分析, 对高脂饮食诱导下金黄地鼠粪便和空肠、回肠、盲肠、结肠四段肠道内容物的代谢轮廓进行研究, 观察代谢物分子水平上高脂饮食对肠道代谢的影响, 为肠道菌群在高脂血症疾病发展过程中起到的作用提供分子基础, 为高脂血症的药物研发及临床治疗提供了有用信息及新的研究思路。
材料与方法实验动物 24只雄性叙利亚金黄地鼠(4~5周龄), 由北京维通利华实验动物技术有限公司提供(合格证号: SCXK北京2009-0008), 饲养于中国医学科学院药物研究所动物实验中心屏障环境设施中。
试剂 羧甲基纤维素钠(carboxymethyl cellulose sodium, CMC-Na, 批号F20031117)、NaH2PO4·2H2O (分析纯)和K2HPO4·3H2O (分析纯)均购自上海市国药集团化学试剂有限公司。3-(三甲基甲硅烷硅基)-1-丙磺酸钠[3-(trimethylsilyl)-1-propanesulfonic acid-d6 sodium salt, DSS]为青岛腾龙微波有限公司产品。
试剂盒 甘油三酯检测试剂盒(批号: 805RCK)、总胆固醇检测试剂盒(批号: 803RJJ)均购自日本积水医疗株式会社, 规格为通用型, 2~8 ℃保存; PEG6000沉淀剂, 批号: BCBK6550V, Sigma公司产品。
实验仪器 多用途低温高速离心机, 德国Eppendorf公司产品; SeperateTM Max 190酶标仪, Molecular Devices公司产品; 超声破碎仪, 中国赛飞公司产品; Bruker AVANCE Ⅲ-500超导傅立叶变化核磁共振波谱仪, 瑞士布鲁克公司产品。
高血脂地鼠模型建立 本实验采用高脂饲料喂饲法诱导金黄地鼠高脂模型的形成。24只金黄地鼠适应性饲养一周后, 空白对照组(12只), 每日给予基础饲料, 高脂模型组(12只)给予高脂饲料(胆固醇0.3%, 猪油14.5%, 基础饲料85.2%)喂养, 诱导高血脂症模型。建立高脂模型时, 动物自由摄食饮水, 饲养温度为20~24 ℃, 空气湿度为35%~55%, 光照12 h, 黑暗12 h。
血清生化指标检测 血清TC、TG、HDL-C和LDL-C测定按照试剂盒说明书进行。
粪便和肠道内容物预处理与1H NMR测定 称取新鲜粪便和肠道内容物(空肠、回肠、盲肠和结肠内容物)约200 mg于离心管中, 编号并记录重量(其中由于结肠内容物样本量不足, 仅测试12例样本, 其中对照组6例, 模型组6例)。向离心管中加入磷酸盐缓冲液(100% H2O, 0.15 mol·L-1 NaH2PO4/K2HPO4, pH 7.4) 1 mL, 涡旋样本1 min。之后将样本于冰水浴中, 超声破碎, 超声4 s, 间歇4 s, 如此循环16次。破碎后于4 ℃、10 000×g离心15 min。取上清450 μL, 加入磷酸盐缓冲液(100% D2O, 0.15 mol·L-1 NaH2PO4/K2HPO4, 5 mmol·L-1 DSS, pH 7.4) 50 μL, 混匀, 转移至5 mm核磁管, 于25 ℃进行1D NOESY 1H NMR测试。
1H NMR图谱处理与多元统计分析 对NMR分析所得谱图进行傅立叶变换、调相、调基线后, 定标。核磁波谱数据调用AMIX (analysis of mixtures software v. 3.0, Bruker Biospin)软件中的程序处理。肠道内容物1D NOESY 1H NMR图谱按0.68~9.5 ppm (除去水峰4.62~5.18 ppm, DSS峰1.7~1.8和2.84~2.94 ppm)的范围分段积分。其中每段为0.04 ppm。将积分数据归一化之后, 将积分值进行中心化和比例换算, 用SIMCA-P12.0软件包(瑞典, Umetrics AB, Umeå)进行OPLS-DA统计分析。
结果 1 高脂金黄地鼠模型血脂水平的改变分别于2周、4周和8周时测量正常对照组与高脂饮食组血脂水平, 由表 1可知各时间点高脂饮食组血清中TC、TG、LDL-C水平均显著高于空白对照组, 且具有统计学差异(P<0.05); HDL-C在2周和4周时显著增加, 但在8周时降低, 且差异无统计学意义(P>0.05)。其中TC、TG、LDL-C的升高表明高脂饮食诱导地鼠血脂积累, 高脂血症金黄地鼠模型的建立是成功的。
| Table 1 Serum biochemistry test results from control and high-fat diet (HFD) groups of hamsters. TG: Triglyceride; TC: Total cholesterol; LDL-C: Low density lipoprotein cholesterol; HDL-C: High density lipoprotein cholesterol. n = 12. #P < 0.05, ##P < 0.01, ###P < 0.001 vs control |
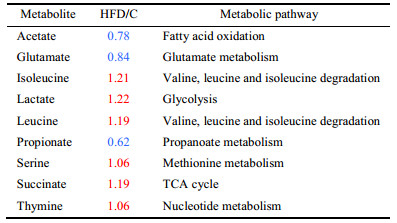
首先采用OPLS-DA模式识别对高脂饮食组和正常组粪便代谢轮廓分析进行深入系统的比较。由图 1a得分图可知, 高脂饮食诱导后, 高脂模型组和正常组粪便的代谢轮廓产生明显区分R2X = 0.713, R2Y = 0.97, Q2 = 0.801; CV-ANOVA评估, P<0.05 (P = 0.002)。利用Chenomx NMR Suite (V. 7.5, Chenomx, Edomonton, Canada)软件进行鉴定, 并结合文献[12]进行归属, 从图 1b载荷图可以得到高脂血症金黄地鼠与正常对照组的差异代谢物(筛选VIP>1且独立样本t检验中P<0.05的代谢产物[13])。作为模型组粪便代谢组相对于正常组升高的代谢物有乳酸、琥珀酸、亮氨酸等6种, 降低的代谢物有乙酸、丙酸、谷氨酸3种, 具体代谢物变化见表 2。

|
Figure 1 OPLS-DA scores plot (a) and loadings plot (b) derived from 1H NMR spectra of feces. HFD: High-fat diet; C: Control. n = 12 |
| Table 2 Differential metabolites of feces, metabolic pathways, as well as fold changes between control and HFD group. The red indicated the HFD-induced significant elevations whereas the blue indicated decreases. n = 12 |
与粪便代谢轮廓分析相似, 采用OPLS-DA模式识别分析分别对高脂饮食组和正常组的空肠、回肠、盲肠和结肠内容物代谢轮廓分析进行比较(图 2)。各个肠道内容物模型组和正常对照组之间发生明显改变并具有统计学意义(P<0.05, 表 3)。观察高脂模型组与正常对照组的差异代谢物, 其中高脂模型组空肠内容物与正常对照组相比丙氨酸、苯丙氨酸、酪氨酸和苏氨酸4种代谢物水平显著升高; 回肠内容物苯丙氨酸水平显著降低, 琥珀酸水平显著升高。盲肠内容物在高脂饮食8周后胆碱水平显著升高, 葡萄糖、酪氨酸、尿嘧啶水平显著降低; 结肠中乙酸、丙氨酸、次黄嘌呤等7种代谢物水平降低, 2-氧代异己酸水平升高(图 3)。其中空肠和回肠属于小肠, 盲肠和结肠属于大肠, 小肠的代谢物改变明显少于大肠。

|
Figure 2 OPLS-DA scores plot (left) and loadings plot (right) derived from 1H NMR spectra of intestinal contents. a: Jejunum; b: Ileum; c: Cecum; d: Colon. For a, b and c, n = 12; For d, n = 6 |
| Table 3 Basic parameters of OPLS-DA models |

|
Figure 3 Different metabolites in jejunum (a), ileum (b), cecum (c) and colon (d) contents. *P < 0.05, **P < 0.01, ***P < 0.001 vs control. n = 12 in a, b and c; n = 6 in d |
脂代谢紊乱被认为是心血管疾病的病理基础, 其中以高脂血症为代表的临床病症是诱发动脉粥样硬化、冠心病、高血压、糖尿病、胰腺炎、癌症等多种重大疾病的主要因素之一。临床上对其诊断依据主要是血清TC、TG、LDL-C、HDL-C和一些影像学指标, 这些指标受限于对该类疾病在中晚期的描述, 难于对疾病发生发展进程进行全面的解释及动态跟踪。一旦确诊, 此类疾病其实早已形成, 甚至已造成难以逆转的器质性损害。因此, 为了更全面地了解脂代谢紊乱的发病机制, 开发新方法越来越被需要。越来越多的研究表明肠道菌群在脂代谢紊乱疾病的发生发展过程中起着重要作用[14-16]。同时, 针对调节肠道菌群, 改善脂代谢紊乱状态, 从而治疗由此引发的一系列心血管疾病, 已成为调血脂药物研发的一个热点[17], 而肠道菌群的生存及改变与其在宿主中的环境密切相关, 宿主肠道中的内源性代谢物变化可为肠道菌群变化提供直接的证据。
在之前的研究中高脂饮食诱导的高脂血症模型组血清中TC、TG、LDL-C水平显著升高, 表明模型的成功建立[18], 其中血清中TC和LDL-C水平的升高是动脉粥样硬化发展的重要危险因素[4]。在上述的研究结果中, 相比对照组, 模型组从2周开始TC、TG、LDL-C水平显著升高, 此种趋势一直延续至8周实验结束, 表明高脂血症模型建立成功, 同时表明模型组的脂代谢发生紊乱, 进而引起多种生物介质(粪便、肠道内容物)中脂肪酸代谢、氨基酸代谢、胆碱代谢等多条代谢途径发生紊乱。
本研究中高脂饮食诱导8周后粪便中短链脂肪酸(SCFAs)降低, SCFAs有许多生物学功能, 它们能作为结肠上皮细胞的能源物质, 可以促进细胞增殖和分化, 作为胞内或胞外pH指示剂等[19]。最近的研究还表明SCFAs能激活孤儿G蛋白偶联受体[20], 抑制脂肪细胞中的胰岛素信号通路, 从而抑制脂肪组织中的脂肪积累[21]。此外有研究表明, 由于粪便中含有乳酸代谢的菌群, 正常情况下乳酸的含量通常较低, 然而当发生急性期溃疡性结肠炎时, 乳酸由发炎的黏膜细胞分泌出来, 患者粪便中通常含有较高水平的乳酸[22, 23]。在本研究中HFD干预第8周时粪便中乳酸水平升高, 进一步说明高脂血症形成过程中有炎症发生。
此外还观察到高脂饮食干预造成金黄地鼠空肠内容物中苯丙氨酸、酪氨酸、苏氨酸含量上升。有研究表明, 肠道菌群失调会导致炎症的发生, 从而导致肠道中氨基酸因吸收不良而含量增加[24]。针对本研究中盲肠和结肠内容物中发生显著变化的代谢物分析可知, 模型组中由肠道菌群降解产生的单糖, 如葡萄糖含量降低, 由于哺乳动物自身缺乏降解饮食中的膳食纤维相关的特殊酶, 哺乳动物自身难以消化这些纤维, 所以肠道中的单糖一般来源于食物中半纤维素在肠道菌群的作用下降解的产物[25, 26]。如大肠中的厌氧菌, 以拟杆菌为代表, 能分解多种糖苷键。这些单糖对某些肠道菌群来说是重要的能源物质, 单糖水平的变化表明了肠道菌群发生紊乱。同时, 在本研究中还观察到盲肠和结肠内容物中尿嘧啶水平降低, 结肠内容物中次黄嘌呤下降。尿嘧啶和次黄嘌呤是嘧啶和嘌呤衍生物。研究表明, 尿嘧啶和次黄嘌呤水平的下降表明肠道中菌群数量的降低[27]。这表明高脂饮食导致的肠道菌群失调还会造成核苷酸代谢紊乱。结肠内容物模型组中苯丙氨酸、酪氨酸水平降低与尿嘧啶和次黄嘌呤的变化一致, 都表示肠道中总的菌群数量降低。另一方面, 酪氨酸和苯丙氨酸属于芳香族氨基酸, 可由肠道菌群代谢生成对羟基苯乙酸, 随后生成对甲酚[28], 酪氨酸和苯丙氨酸水平的降低还表明在高脂饮食的干预下, 参与这一过程的特定的肠道菌群数量的增加或者活性加强。
本研究中对比高脂饮食对小肠和大肠的影响, 其中四段肠道内容物中空肠回肠属于小肠, 盲肠回肠属于大肠, 本研究发现小肠中发生显著变化的代谢物种类少于大肠, 说明高脂饮食对小肠的影响弱于大肠。推测其原因, 可能是由于小肠中富含营养物质, 可为菌群提供的营养成分较充足, 且大部分细菌依靠胆盐水解酶水解结合胆盐, 并对分离下来的甘氨酸、牛磺酸加以利用, 从而生长繁殖, 故受饮食结构的影响较小。而大肠中的营养成分主要为未被小肠消化吸收的碳水化合物, 提供给菌群的营养成分相对较少, 因此大肠微生态系统受饮食结构的影响较大且持久, 进一步深究其原因主要为食物中的脂类增多, 使大肠中菌群的养料来源减少、生存环境恶化; 此外脂代谢过程中也会产生部分可损害肠黏膜屏障的副产物, 如次级胆酸、硫化氢等, 导致黏膜产生炎性反应等不良变化, 破坏菌群赖以生存的微环境[29, 30]。
综上所述, 本研究以1H NMR代谢组学技术结合多元统计分析方法对高脂血症金黄地鼠模型粪便和肠道内容物的代谢轮廓进行研究, 分别在粪便、空肠、回肠、盲肠和结肠中鉴定出了高脂模型组和对照组之间差异代谢物, 包括氨基酸代谢物、蛋白质腐败产物、核苷酸代谢物, 胆碱代谢物, 使得肠道菌群和高脂血症之间的紧密关系再一次被证实, 为后续肠道菌群的研究提供了分子基础, 为高脂血症的药物研发及临床治疗提供了有用的信息及新的研究思路。
| [1] | Shen YH, Jin X, Zhao Y, et al. The hypolipidemic and liver protective effect of (Z)-N-(2-hydroxyethyl) docos-13-enamide on hyperlipidemic golden hamsters[J]. Acta Pharm Sin (药学学报), 2012, 47: 1252–1256. |
| [2] | Li ZY, Ding LL, Li JM, et al. 1H NMR and MS based metabolomics study of the intervention effect of curcumin on hyperlipidemia mice induced by high-fat diet[J]. PLoS One, 2015, 10: e0120950. DOI:10.1371/journal.pone.0120950 |
| [3] | Miao H, Chen H, Pei S, et al. Plasma lipidomics reveal profound perturbation of glycerophospholipids, fatty acids, and sphingolipids in diet-induced hyperlipidemia[J]. Chem Biol Interact, 2015, 228: 79–87. DOI:10.1016/j.cbi.2015.01.023 |
| [4] | Song X, Wang J, Wang P, et al. 1H NMR-based metabolomics approach to evaluate the effect of Xue-Fu-Zhu-Yu decoction on hyperlipidemia rats induced by high-fat diet[J]. J Pharm Biomed Anal, 2013, 78-79: 202–210. DOI:10.1016/j.jpba.2013.02.014 |
| [5] | Demyen M, Alkhalloufi K, Pyrsopoulos NT. Lipid-lowering agents and hepatotoxicity[J]. Clin Liver Dis, 2013, 17: 699–714. DOI:10.1016/j.cld.2013.07.016 |
| [6] | Martin FP, Wang Y, Yap IK, et al. Topographical variation in murine intestinal metabolic profiles in relation to microbiome speciation and functional ecological activity[J]. J Proteome Res, 2009, 8: 3464–3474. DOI:10.1021/pr900099x |
| [7] | Wang Y, Holmes E, Comelli EM, et al. Topographical variation in metabolic signatures of human gastrointestinal biopsies revealed by high-resolution magic-angle spinning 1H NMR spectroscopy[J]. J Proteome Res, 2007, 6: 3944–3951. DOI:10.1021/pr0702565 |
| [8] | Wang Y, Tang H, Holmes E, et al. Biochemical characterization of rat intestine development using high resolution magic-anglespinning 1H NMR spectroscopy and multivariate data analysis[J]. J Proteome Res, 2005, 4: 1324–1329. DOI:10.1021/pr050032r |
| [9] | Martin FP, Wang Y, Sprenger N, et al. Effects of probiotic Lactobacillus paracasei treatment on the host gut tissue metabolic profiles probed via magic-angle-spinning NMR spectroscopy[J]. J Proteome Res, 2007, 6: 1471–1481. DOI:10.1021/pr060596a |
| [10] | Dumas M, Barton RH, Toye A, et al. Metabolic profiling reveals a contribution of gut microbiota to fatty liver phenotype in insulin-resistant mice[J]. Proc Natl Acad Sci U S A, 2006, 103: 12511–12516. DOI:10.1073/pnas.0601056103 |
| [11] | Jove M, Pamplona R, Prat J, et al. Atherosclerosis prevention by nutritional factors:a meta-analysis in small animal models[J]. Nutr Metab Cardiovasc Dis, 2013, 23: 84–93. DOI:10.1016/j.numecd.2012.09.006 |
| [12] | Tian Y, Zhang L, Wang YL, et al. Age-related topographical metabolic signatures for the rat gastrointestinal contents[J]. J Proteome Res, 2012, 11: 1397–1411. DOI:10.1021/pr2011507 |
| [13] | Xia XT, Sun N, Liu CC, et al. Discovering potential biomarkers of depression and drug intervention of paroxetine based on 1H NMR metabolomics[J]. Acta Pharm Sin (药学学报), 2016, 51: 595–599. |
| [14] | Dunne C. Adaptation of bacteria to the intestinal niche:probiotics and gut disorder[J]. Inflamm Bowel Dis, 2001, 7: 136–145. DOI:10.1097/00054725-200105000-00010 |
| [15] | Turnbaugh P, Ley R, Mahowald M, et al. An obesityassociated gut microbiome with increased capacity for energy harvest[J]. Nature, 2006, 444: 1027–1031. DOI:10.1038/nature05414 |
| [16] | Dethlefsen L, Fall-Ngai M, Relman DA. An ecological and evolutionary perspective on human-microbe mutualism and disease[J]. Nature, 2007, 449: 811–818. DOI:10.1038/nature06245 |
| [17] | Wu H, Esteve E, Tremaroli V, et al. Metformin alters the gut microbiome of individuals with treatment-naive type 2 diabetes, contributing to the therapeutic effects of the drug[J]. Nat Med, 2017, 23: 850–858. DOI:10.1038/nm.4345 |
| [18] | Shi F, Li JL, Yang LQ, et al. Hypolipidemic effect and protection ability of liver-kidney functions of melanin from Lachnum YM226 in high-fat diet fed mice[J]. Food Funct, 2018, 9: 880–889. DOI:10.1039/C7FO01294B |
| [19] | Cook SI, Sellin JH. Review article:short chain fatty acids in health and disease[J]. Aliment Pharmacol Ther, 1998, 12: 499–507. DOI:10.1046/j.1365-2036.1998.00337.x |
| [20] | Gill SR. Metagenomic analysis of the human distal gut microbiome[J]. Science, 2006, 312: 1355–1359. DOI:10.1126/science.1124234 |
| [21] | Kimura I. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43[J]. Nat Commun, 2013, 4: 1829. DOI:10.1038/ncomms2852 |
| [22] | Vernia P, Caprilli R, Latella G, et al. Fecal lactate and ulcerative colitis[J]. Gastroenterology, 1988, 95: 1564–1568. DOI:10.1016/S0016-5085(88)80078-7 |
| [23] | Hove H, Nordgaard-Andersen I, Mortensen PB. Faecal DL-l actate concentration in 100 gastrointestinal patients[J]. Scand J Gastroenterol, 1994, 29: 255–259. DOI:10.3109/00365529409090473 |
| [24] | Marchesi JR, Holmes E, Khan F, et al. Rapid and noninvasive metabonomic characterization of inflammatory bowel disease[J]. J Proteome Res, 2007, 6: 546–551. DOI:10.1021/pr060470d |
| [25] | Cabotaje LM, Lopezguisa JM, Shinnick FL, et al. Neutral sugar composition and gravimetric yield of plant and bacterial fractions of feces[J]. Appl Environ Microbiol, 1990, 56: 1786–1792. |
| [26] | Crittenden R. In vitro fermentation of cereal dietary fibre carbohydrates by probiotic and intestinal bacteria[J]. J Sci Food Agric, 2002, 82: 781–789. DOI:10.1002/jsfa.1095 |
| [27] | Hong YS, Ahn YT, Park JC, et al. 1H NMR based metabonomic assessment of probiotic effects in a colitis mouse model[J]. Arch Pharm Res, 2010, 33: 1091–1101. DOI:10.1007/s12272-010-0716-1 |
| [28] | Lesaffer G, De-Smet R, Belpaire FM, et al. Urinary excretion of the uraemic toxin p-cresol in the rat:contribution of glucuronidation to its metabolization[J]. Nephrol Dial Transplant, 2003, 18: 1299–1306. DOI:10.1093/ndt/gfg107 |
| [29] | Resta SC. Effects of probiotics and commensals on intestinal epithelial physiology:implications for nutrient handing[J]. J Physiol, 2009, 587: 4169–4174. DOI:10.1113/jphysiol.2009.176370 |
| [30] | Attene-Ramos MS, Nava GM, Muellner MG. DNA damage and toxicogenomic analyses of hydrogen sulfide in human intestinal epithelial FHs 74 Int cells[J]. Environ Mol Mutagen, 2010, 51: 304–314. |