 2016, Vol. 51
2016, Vol. 51



新生隐球菌 (Cryptococcus neoformans) 是一种条件性致病真菌[1],其主要通过呼吸道入侵肺部引发肺隐球菌病,再经血脑屏障进一步感染中枢神经系统导致隐球菌脑膜炎[2, 3]。因此,感染者常常同时存在肺部感染和中枢神经系统感染,病情复杂且病死率极高。据统计,全球每年由于隐球菌病死亡的人数超过60万[1]。但临床上抗真菌药物相对匮乏,且现有药物大多存在毒副作用明显、易产生耐药性等问题[4, 5]。随着易感人群 (AIDS患者和器官移植人群等) 的快速增长,寻找抗隐球菌感染更为有效的治疗手段、研发治疗隐球菌病的新型药物已迫在眉睫。
新药研发的历史和经验提示,除了不断发掘和设计新的化合物外,通过新制剂技术来“改良”已有药物往往具有同等的重要性。两性霉素B脂质体 (AmBisome) 作为屈指可数的抗真菌药物新制剂[6],即是重要代表之一。通过微粒递药系统的包载能显著改善许多药物的理化性质及体内行为。与两性霉素B去氧胆酸盐注射液相比,脂质体制剂能明显降低原药的肾毒性并提高疗效[7, 8]。基于递药系统自身特点 (如粒径大小、表面性质等) 以及体内生理环境对其的影响程度,一些微粒递药系统已能表现出在病灶部位及相关组织、脏器的较高蓄积,是被动靶向[9]; 而在此基础上于载体表面修饰目标细胞特定受体的配体,通过受体-配体特异性相互作用能介导载体进一步富集于靶区,被称为主动靶向[10]。主动靶向策略在抗肿瘤和部分脏器疾病研究领域得到广泛应用[11, 12],近年来已逐步拓展到抗病原微生物方面。一些研究表明,借助能识别病原菌的相应抗体、多肽等导向分子的“引导”功能,纳米体系或制剂的抗菌效果能得到显著提高[13, 14]。不同于针对感染所在单一脏器的靶向策略,递药系统若能直接识别病原菌,可能会在抗多脏器感染的治疗中有其特殊优势。
导向分子是实现主动靶向递药的重要物质基础之一,对于抗病原菌感染而言,新导向分子的发现还对构建相应的诊断方法具有潜在价值。多肽因其易于制备、对靶蛋白亲和力高和专一性强、潜在免疫原性低等优点,是介导靶向递药的一类重要且被广泛应用的导向分子[15]。能发挥“引导”功能的多肽分子来源多样,其中,噬菌体展示 (肽库) 是获得多肽导向分子的一种重要技术[16]。除了能针对已知靶蛋白筛选相应多肽配体,噬菌体展示的魅力还在于能够直接开展细胞乃至体内水平的功能多肽筛选, 因而该技术与靶向递药研究结合日趋紧密[17, 18, 19]。基于此,本文拟通过噬菌体展示筛选能特异性结合新生隐球菌的多肽分子,将该多肽修饰于脂质体表面并评价其体外及体内靶向性; 进一步包载模型药物伊曲康唑后初步评价其抗隐球菌肺脑合并感染的药效,以期为抗真菌感染治疗提供一种新的药剂学手段。
材料与方法 药品与试剂噬菌体随机12肽库 (NEB PHD12 #E8100S,美国New England Biolabs公司); M13噬 菌体单链DNA快速提取试剂盒 (Cat#N1013,明日百傲科技有限公司); 伊曲康唑 (ITZ,上海思域化工科技有限公司); 蛋黄卵磷脂 (EPC)、胆固醇 (CHO)、聚乙二醇-二硬脂酰乙醇胺 (mPEG2000-DSPE) (上海艾韦特医药科技有限公司); 马来酰亚胺-聚乙二醇-磷脂酰乙醇胺 (Mal-PEG-DSPE,美国Layson Bio公司); 1,1-(十八烷基)-3,3,3,3'-四甲基吲哚三碳菁碘 (DiR,美国Invitrogen公司); 6-氨基荧光素 (6-FAM,南京森贝伽生物科技),其他试剂均为分析纯。
仪器石英晶体微天平 (qCell T,德国3T analytik公司); 生物质谱仪 (德国布鲁克公司); 微型挤出器 (mini-extruder,Avanti Polar Lipids公司); 激光粒度 分析仪 (Nano ZS,英国马尔文公司); 流式细胞仪 (ACEA Novo Cyte,杭州艾森生物工程有限公司); 荧光倒置显微镜 (IX-73,日本奥林巴斯公司); 小动物活体成像系统 (FX PRO,美国布鲁克公司); 酶标仪 (BIO-RAD Model 680); 高效液相色谱仪 (HPLC,LC-20AT,日本岛津公司)。
实验菌种新型隐球菌H99 (Cryptococcus neoformans) 由美国罗格斯大学薛朝阳教授惠赠。挑取单菌落于酵母膏胨葡萄糖 (yeast extract peptone dextrose,YPD) 培养基中,于30 ℃条件下过夜培养。实验真菌为处于对数生长期的真菌。
实验动物昆明小鼠,雄性,体重 (20 ± 2) g,许可证号: SCXK-(军)2012-0011,购自重庆腾鑫生物技术有限公司。实验小鼠由西南大学药学院实验动物 中心进行屏障系统SPF级动物房饲养,自由饮食和饮水。
噬菌体展示筛选及分析隐球菌分散于pH 7.4磷酸盐缓冲液 (PBS) 中; 噬菌体库由TBST (50 mmol·L-1 Tris-HCl,150 mmol·L-1 NaCl pH 7.5,0.1% Tween 20) 稀释到1×1010 pfu/mL,两者于室温孵育 2 h。未结合的噬菌体经TBST清洗、离心后除去 (弃去上清),重复5次。用酸洗脱法 (50 mmol·L-1 Glycine-HCl,pH 2.0) 将结合的噬菌体洗脱下来,加入适量Na2HPO4缓冲液 (pH 7.5) 中和酸性; 洗脱下来的噬菌体加入对数期大肠杆菌ER2738菌体振荡 4 h进行扩增,并用于第2轮筛选。同样的方法筛选 3轮后,回收噬菌体洗脱产物。每轮回收得到的噬菌体通过双层琼脂平板法 (LB/X-gal/IPTG铺板) 测定其滴度,噬菌体纯化通过PEG沉淀法进行。用M13噬菌体单链DNA快速提取试剂盒对3轮筛选后所得单克隆噬菌体进行DNA提取,琼脂糖凝胶电泳检测所提DNA大小是否正确。选取噬菌体展示文库试剂盒内-96 gIII引物用于自动测序,测序由上海生工公司完成。利用Snapgene Viewer 3.1软件对单克隆噬菌体DNA序列进行分析。所读序列为模板序列的反义链,即序列与模板序列互补。根据遗传密码表可翻译获得展示肽段的氨基酸序列。
多肽与真菌结合的初步考察所有多肽N端均偶联半胱氨酸。常规清洗石英晶体微天平 (Quartz Crystal Microbalance,QCM) 后通入PBS,待芯片振动频率稳定后,通入1 mg·mL-1多肽水溶液使多肽与金芯片键合,再通入PBS除去过量未与芯片键合的多肽,20 min后频率达到稳定。将PBS置换成浓度为1×107 CFU/mL菌液,120 min后记录芯片振动频率的变化值 (△F),以上流速均为50 µL·min-1。常规清洗仪器后,将隐球菌菌液替换为液体培养基 (YPD),重复以上实验,作为空白对照组。
功能膜材料的制备与表征合成多肽偶联的聚乙二醇-磷脂酰乙醇胺: 将MAL-PEG-DSPE与N 端巯基化的多肽按照摩尔比1∶1.2分别溶解于二甲基甲酰胺 (DMF) 和PBS中,涡旋混合均匀,磁力搅拌器37 ℃匀速搅拌24 h,采用薄层色谱 (thin layer chromatography,TLC) 监测反应进行程度。反应达到终点时将反应物对纯水透析 (截留分子质量3.5 kDa),除去过量未反应的多肽和DMF。冷冻干燥后于 -20 ℃储存备用。采用生物质谱仪 (MALDI-TOF MS) 分别测定合成终产物P-PEG-DSPE和Mal-PEG-DSPE的分子量,以证明NNH肽 (序列NNHREPPDHRTS) 与Mal-PEG-DSPE聚合成功。
脂质体的制备蛋黄卵磷脂 (EPC)、胆固醇 (CHO)、高分子材料 (mPEG2000-DSPE) 和功能膜材料按摩尔比62∶36∶1∶1,药脂比为1∶25,溶于适量氯仿,减压除去有机溶剂并旋转成膜,以pH 7.4 PBS于37 ℃水化,经微型挤出器依次通过200 nm 和100 nm膜,即得带有导向分子的伊曲康唑脂质体 (P-LIP-ITZ)。普通伊曲康唑脂质体的制备与上述方法相似,只是制备时不加功能膜材料。按同样方法制备包载不同荧光探针的脂质体用于后续表征实验 (以6-FAM为荧光探针分别制备P-LIP-FAM和LIP-FAM,用于流式细胞检测; 以DiR为荧光探针分别制备P- LIP-DiR和LIP-DiR,用于荧光成像检测) 。
脂质体的表征对所得脂质体进行粒径和多分散度 (PDI) 测定。采用HPLC法测定药物包封率。绘制伊曲康唑在液相色谱条件 (色谱柱: ODS-SP C18柱150 mm × 5 mm,5 µm; 流动相: 水17%,甲醇83%; 流速: 1 mL·min-1; 检测波长: 262 nm; 柱温: 25 ℃) 下的标准曲线。测定伊曲康唑脂质体包封率: 取载药后未经纯化的伊曲康唑脂质体两份,一份经葡聚糖凝胶柱纯化后收集,另一份不作其他处理仅稀释到相同体积,均加入5倍体积甲醇破乳24 h,采用HPLC法测定总的伊曲康唑浓度 (C0) 以及脂质体中伊曲康唑的浓度 (C)。按以下公式计算包封率 (EE)。
EE = C / C0 × 100%
体外隐球菌与脂质体的结合将H99隐球菌接种于YPD液体培养基中过夜培养,处于对数生长期的隐球菌用PBS洗涤2次后重悬于新鲜的YPD,终浓度为1×107 CFU/mL。制备包载相同含量6-FAM的脂质体 (P-LIP-FAM、LIP-FAM) 并分别与真菌悬液混合,设立空白真菌对照组,于30 ℃条件下培养 (1、2和4 h),各组于设定时间点取样后PBS洗涤2次,流式细胞仪测定脂质体对真菌的结合情况 (6-FAM,激发波长为495 nm,发射波长为519 nm)。
隐球菌感染模型建立将H99隐球菌接种于YPD液体培养基中30 ℃培养过夜,处于对数生长期的隐球菌使用PBS洗涤2次重悬于灭菌生理盐水,终浓度为1×107 CFU/mL。小鼠尾静脉注射菌液200 μL,观察小鼠的存活情况。在出现感染症状后,无菌环境下使用毛细管抽取小鼠脑脊液并解剖取出该小鼠的脑和肺,用生理盐水清洗,滤纸吸干并称重后置于EP管中 ,加入生理盐水1 mL,匀浆。将脑脊液稀释10倍,组织匀浆液稀释100倍和10 000倍,分别吸取各样品25 μL涂布于YPD固体培养基,30 ℃培养48 h,观察菌落的生长情况。
体内靶向性考察分别制备粒径相当的载近红外荧光探针 (DiR 100 μg·mL-1) 脂质体P-LIP-DiR和LIP-DiR。8只感染模型小鼠随机分为两组,尾静脉注射P-LIP-DiR和LIP-DiR各100 μL; 另取健康小鼠4只于尾静脉注射P-LIP-DiR 100 μL。各组于1、2、4和8 h分别解剖取出脑、心、肺、肝、脾和肾脏进行荧光成像检测 (DiR,激发波长为720 nm,发射波长为790 nm)。
组织分布实验分别制备粒径相当的载药 (ITZ) 靶向脂质体 (P-LIP-ITZ) 和普通脂质体 (LIP-ITZ)。24只感染模型小鼠随机分为两组,分别尾静脉注射给予P-LIP-ITZ和LIP-ITZ; 另取12只健康小鼠尾静脉注射P-LIP-ITZ。各组小鼠ITZ给药量均为12 mg·kg-1,于1、2、4和8 h分别解剖,每组每个时间点取3只小鼠,分别取脑、肺、肝组织,用生理盐水清洗,滤纸吸干并称重,置于10 mL EP管中,加入生理盐水0.5 mL,匀浆,再加入1 mL甲醇涡旋振荡萃取ITZ,4 000 r·min-1离心10 min后取上清液于4 mL离心管中,氮气吹干,取流动相200 µL复溶,进行液相检测。
体外药效学实验微板稀释法考察ITZ溶液(溶于5% DMSO) 及两种脂质体制剂 (LIP-ITZ和P- LIP-ITZ) 的抑菌能力[20]。分别将上述3种药液用YPD培养基稀释到终质量浓度为2 µg·mL-1,吸取100 µL置96孔板的第1个孔,加入YPD 100 µL混匀,从 中吸取100 µL加入第2个孔,按照此类2倍稀释 法进行稀释,形成药物质量浓度梯度为1、0.5、0.25、0.125、0.063、0.032和0.016 µg·mL− ;1,每孔内加入隐球菌菌液 (1×106 CFU/mL) 100 µL。另设两组对照,一组加菌不加药,观察隐球菌在实验培养条件下能否正常生长; 另一组加药不加菌,观察在实验操作过程中是否存在污染。接种的96孔板置于30 ℃培养箱过夜培养。48 h后观察并测定每孔在630 nm 波长下的吸光度 (OD) 值,判断各含药组的抑菌能力。
体内药效学实验将44只感染模型小鼠随机分为4组,分别设为非治疗组、伊曲康唑包合物注射液 (CD-ITZ) 组、LIP-ITZ组和P-LIP-ITZ组。在小鼠出现明显感染症状后 (感染第3天) 给药1次,相应制剂经尾静脉注射200 μL (ITZ 8 mg·kg-1),饲养观察各组小鼠在15天内的存活情况。于感染后第6天,每组各取小鼠1只解剖获得脑、肺、肝3种组织,生理盐水洗净后置于4% 福尔马林中浸泡过夜,石蜡切片进行过碘酸雪夫染色 (PAS) 实验,观察各组织中隐球菌的分布情况。
数据分析应用SPSS17.0软件对组织分布实验中各组织的含药量及体内药效实验中小鼠生存时间进行统计学分析,P < 0.05为具有显著性差异。
结果与讨论 1功能多肽筛选与初步考察经3轮筛选后,得到重现性较强的两条序列,分别为: NNHREPPDHRTS和GEVGEQEKARVG (GEV)。拟通过QCM来初步评价多肽与真菌的相互作用。QCM测定原理为,当通入物质吸附在芯片表面时会引起芯片振动频率的变化,振动频率减少则提示芯片上吸附的质量有所增加,测量精度可达纳克级。已有文献[21]报道使用QCM考察细菌与抗菌肽的相互作用能力。这里使用QCM测定在加入菌液或培养基后芯片振动频率的变化量。若隐球菌能与多肽结合,则当越多的隐球菌通过多肽停留于金芯片时,芯片的振动频率变化值越大 (减少量越多)。由 (图 1) 所示,多肽NNH对隐球菌结合明显,显著高于另一条多肽GEV。作者还合成了一条乱序肽 (RNPNHRTEDSHP,氨基酸组成完全一致但排列随机打乱),经测定,乱序肽对隐球菌无明显结合。而通入YPD时均未引起上述含肽芯片振动频率的明显变化。实验结果初步说明,NNH肽能特异性结合真菌,将作为潜在的真菌靶向导向分子用于后续研究。

|
Figure 1 Quartz crystal microbalance (QCM) detection of peptide binding to Cryptococcus after injection of 1×107 CFU/mL Cryptococcus into a peptide-immobilized chip. The medium served as controls. n = 3,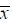 ± s. NNH: Peptide sequences,NNHREPPDHRTS; GEV: Peptide sequences,GEVGEQEKARVG ± s. NNH: Peptide sequences,NNHREPPDHRTS; GEV: Peptide sequences,GEVGEQEKARVG
|
功能膜材料的合成原理是利用马来酰亚胺与 活性巯基发生Michael反应形成稳定的硫醚键。该 反应在中性pH环境下具有较高的反应活性,条件 温和且易于键合,常用于构建多肽偶联的靶向递药系统。合成所用多肽的量略过量于Mal-PEG-DSPE,通过TLC法检测提示反应已完全。MALDI-TOF MS表征如图 2,质谱峰所示Mal-PEG-DSPE分子质量为4 206.245 Da,P-PEG-DSPE的分子质量为 5 781.236 Da,聚合反应前后的分子质量之差与NNH肽分子质量的理论值 (1 562 Da) 几乎一致,结果表明该目标产物已成功合成。

|
Figure 2 MALDI-TOF mass spectrometry analysis of Mal- PEG-DSPE and P-PEG-DSPE. Mal-PEG-DSPE: Maleimide- PEG-DSPE; P-PEG-DSPE: Peptide-PEG-DSPE |
伊曲康唑是一种三氮唑类抗真菌药,其能抑制真菌细胞色素P450,对菌体产生破坏杀灭真菌。市面上现有的伊曲康唑剂型主要有口服胶囊、口服液和注射液 (包合物),因其生物利用度不高只作为二线抗真菌药物使用。伊曲康唑脂溶性强,可以稳定地嵌入脂质体的疏水区,本文选择伊曲康唑作为模型药物。
所得载药脂质体的粒径均较小,平均粒径在90 nm左右 (图 3),分散均匀,包封率高,提示表面经导向分子修饰后不影响载药脂质体的基本性质: P-LIP-ITZ的平均粒径为88.25 ± 2.43 nm,分布较均匀,PDI为0.143 ± 0.016,包封率达 (88.05 ± 0.25) %; LIP-ITZ 的平均粒径为86.33 ± 3.84 nm,PDI为0.115 ± 0.012,包封率达 (89.36 ± 0.31) %。

|
Figure 3 Transmission electron micrograph of LIP-ITZ and P-LIP-ITZ (A,bar represents 100 nm); partical size distribution of LIP-ITZ and P-LIP-ITZ (B). LIP-ITZ: Itraconazole liposome; P-LIP-ITZ: Peptide modified itraconazole liposome |
尾静脉注射菌液是建立小鼠隐球菌感染模型的方法之一,其优点是隐球菌通过尾静脉注射容易定量,误差小,不同批次小鼠造模后感染症状表现更具有一致性[22]。图 4所示,分别为正常小鼠和感染小鼠的脑脊液 (稀释10倍)、脑及肺部 (匀浆后分别稀释 100倍和10 000倍) 真菌负荷量。正常小鼠组各样本于YPD 30 ℃培养48 h后,未见隐球菌生长,而将感染小鼠组各样本于YPD同样条件下培养后,有一定数量的隐球菌菌落出现,表明模型小鼠的脑脊液、脑和肺部均存在隐球菌,提示小鼠隐球菌合并感染模型构建成功。

|
Figure 4 The number of fungal colonies on yeast extract peptone dextrose medium (YPD) plates. Samples were collected from cerebrospinal fluid (CSF,diluted 10-folds),brain and lung homogenates (diluted 100-folds and 10000-folds) of infected group and normal group,respectively |
脂质体对真菌的结合实验 (流式细胞术) 结果如图 5所示,经P-LIP-FAM处理后的H99隐球菌荧光素量显著高于LIP-FAM组,在1 h时两者荧光强度相差5.72倍。表面经导向分子修饰的脂质体其真菌结合量显著高于未经修饰的脂质体,提示导向分子能有效提高隐球菌对脂质体的摄取。随着时间推移,两组的真菌荧光强度均有所提高,但P-LIP-FAM组的荧光强度始终强于LIP-FAM组。

|
Figure 5 Invitro fungi binding studies. Flow cytometry analysis of either peptide-modified or unmodified FAM-loaded liposomes. a: PBS; b: LIP-FAM; c: P-LIP-FAM. FAM: Fluoresceinamine isomer II; LIP-FAM: FAM liposome; P-LIP-FAM: Peptide modified FAM liposome |
荧光成像实验结果 (图 6) 显示,在各个时间点,经导向分子修饰的脂质体在模型小鼠脑部与肺部的蓄积程度均显著高于未经修饰的脂质体,而在正常小鼠体内,P-LIP-DiR在脑部与肺部的蓄积量不明显,这初步说明导向分子本身不具有脑或肺部的靶向性,是通过主动识别隐球菌从而介导脂质体富集在富含病菌的病灶部位。图 6B还显示,未修饰的脂质体在模型小鼠脑部有一定程度的蓄积 (相对于正常小鼠),这可能是由于新生隐球菌对血脑屏障造成一定程度的破坏,破坏程度与感染的恶性程度相关[23, 24, 25]。此外, 一些黏附于血脑屏障表面的病原菌也可能成为脂质体识别的靶标。

|
Figure 6 Representative ex vivo images (A) and the semi- quantitative analysis (B)of mice 1,2,4 and 8 h after injection of DiR-labeled liposomes: infected mice treated with P-LIP-DiR (a),infected mice treated with LIP-DiR (b),normal mice treated with P-LIP-DiR (c). 1: Brain; 2: Heart; 3: Lung; 4: Liver; 5: Spleen; 6: Kidney |
将ITZ包载于脂质体,小鼠尾静脉给药后,体 内组织分布实验结果与荧光成像实验有相近的趋势(图 7)。P-LIP-ITZ在模型小鼠肺部、脑部的药物含量显著高于普通脂质体组,给药2 h后两组肺部的含药量相差13.75倍(P < 0.001),而4 h时两组脑部含药量相差2.91倍 (P < 0.01)。

|
Figure 7 Invivo tissue distribution of itraconazole in brain,lung and liver after intravenous injection of different formulations (dose 12 mg·kg-1) at 1,2,4 and 8 h. n = 3,± s. **P < 0.01,***P < 0.001
|
微板稀释法中,使用酶标仪在630 nm波长下检测各孔OD值,OD值与真菌浓度呈正相关。由图 8可见,在相同的ITZ浓度下,P-LIP-ITZ对真菌生长的抑制显著强于LIP-ITZ,初步显示出靶向脂质体的抑菌潜力。在8 mg·kg-1较低剂量及单次给药时 (图 9),P-LIP-ITZ平均生存时间为9天,而LIP-ITZ (平均 生存时间6天) 和CD-ITZ (平均生存时间5天) 对 模型小鼠治疗作用不明显,在15天的观察期内未显著提高小鼠的存活率 (未治疗组的小鼠平均生存时间为5天)。分别取各组小鼠的相关脏器进行组织切片并经PAS染色 (图 10),模型小鼠肺、肝和脑等主要脏器明显可见大量菌体 (箭头所指红色圆点)。而经P-LIP-ITZ单次给药后,感染第6天时各相关脏器的病菌侵袭已得到较好地控制。相比而言,LIP-ITZ组和CD-ITZ组的病菌感染仍较为明显。

|
Figure 8 The in vitro dose-response inhibition curve of fungal growth by treatment with different ITZ formulations |

|
Figure 9 Survival curve of different treatment groups during the observation period. Mice in cyclodextrin-itraconazole (CD- ITZ),LIP-ITZ and P-LIP-ITZ groups received a single dose of itraconazole (8 mg·kg-1). n = 10,± s. P < 0.05
|

|
Figure 10 The histopathological assessment of major organs (lung,brain and liver) of the Cryptococcus infectious mice treated with CD-ITZ,LIP-ITZ,P-LIP-ITZ at a dose of 8 mg·kg-1. Healthy mice served as normal. Periodic acid-Schiff (PAS) stains were performed to highlight the fungi distribution (yellow arrow) |
在观察期内,即使是靶向脂质体治疗组,模型小鼠也多数死亡 (感染6天后),这与给药剂量、给药次数及造模时的感染严重程度均存在密切关系,但也提示上述导向分子及脂质体制剂均有待进一步优化。此外,易复发本身也是隐球菌感染的一个重要特征。因此,值得探索的研究内容是如何将器官靶向与主动靶向隐球菌的策略有机结合[26, 27],以实现更为高效和“精准”的递药,进一步提高抗菌疗效的同时也降低药物对感染病灶周围正常组织的不利影响。
结论本研究基于噬菌体展示技术成功筛选出能特异性结合新生隐球菌的多肽,该多肽能有效介导脂质体对体内外病原菌的特异性识别; 在此基础上构建的主动靶向脂质体制剂能显著提高所包载伊曲康唑的治疗效果。上述研究结果有望为克服隐球菌病及其他真菌治疗当前面临困境 (治疗效果有限、感染反复发作、易产生耐药性等) 提供有益的参考。
| [1] | Idnurm A, Bahn YS, Nielsen K, et al. Deciphering the model pathogenic fungus Cryptococcus neoformans[J]. Nat Rev Microbiol, 2005, 3:753-764. |
| [2] | Liu TB, Perlin DS, Xue CY. Molecular mechanisms of cryptococcal meningitis[J]. Virulence, 2012, 3:173-181. |
| [3] | Leongson K, Cousineau-Cote V, Goupil M, et al. Altered immune response differentially enhances susceptibility to Cryptococcus neoformans and Cryptococcus gattii infection in mice expressing the HIV-1 transgene[J]. Infect Immun, 2013, 81:1100-1113. |
| [4] | Saag MS, Powderly WG, Cloud GA, et al. Comparison of amphotericin B with fluconazole in the treatment of acute AIDS-associated cryptococcal meningitis. The NIAID Mycoses Study Group and the AIDS Clinical Trials Group[J]. N Engl J Med, 1992, 326:83-89. |
| [5] | Kligman AM, Lewis FS. In vitro and in vivo activity of candicidin on pathogenic fungi[J]. Exp Biol Med, 1953, 82:399-404. |
| [6] | Tiphine M, Letscher-Bru V, Herbrecht R. Amphotericin B and its new formulations:pharmacologic characteristics, clinical efficacy, and tolerability[J]. Transpl Infect Dis, 1999, 1:273-283. |
| [7] | Robinson RF, Nahata MC. A comparative review of conventional and lipid formulations of amphotericin B[J]. J Clin Pharm Ther, 1999, 24:249-257. |
| [8] | Kreuter J. Drug delivery to the central nervous system by polymeric nanoparticles:what do we know?[J]. Adv Drug Deliv Rev, 2014, 71:2-14. |
| [9] | Wei X, Chen X, Man Y, et al. Brain tumor-targeted drug delivery strategies[J]. Acta Pharm Sin B, 2014, 4:193-201. |
| [10] | Lammers T, Kiessling F, Hennink WE, et al. Drug targeting to tumors:principles, pitfalls and (pre-) clinical progress[J]. J Control Release, 2012, 161:175-187. |
| [11] | Yan ZL, Wang F, Wen Z, et al. LyP-1-conjugated PEGylated liposomes:a carrier system for targeted therapy of lymphatic metastatic tumor[J]. J Control Release, 2012, 157:118-125. |
| [12] | Takara K, Hatakeyama H, Kibria G, et al. Size-controlled, dual-ligand modified liposomes that target the tumor vasculature show promise for use in drug-resistant cancer therapy[J]. J Control Release, 162:225-232. |
| [13] | Luo Y, Hossain M, Wang C, et al. Targeted nanoparticles for enhanced X-ray radiation killing of multidrug-resistant bacteria[J]. Nanoscale, 2013, 5:687-694. |
| [14] | Millenbaugh NJ, Baskin JB, Desilva MN, et al. Photothermal killing of Staphylococcus aureus using antibody-targeted gold nanoparticles[J]. Int J Nanomed, 2015, 10:1953-1960. |
| [15] | Wang X, Li S, Shi Y, et al. The development of site-specific drug delivery nanocarriers based on receptor mediation[J]. J Control Release, 2014, 193:139-153. |
| [16] | Benhar I. Biotechnological applications of phage and cell display[J]. Biotechnol Adv, 2001, 19:1-33. |
| [17] | Pasqualini R, Ruoslahti E. Organ targeting in vivo using phage display peptide libraries[J]. Nature, 1996, 380:364-366. |
| [18] | Deramchia K, Jacobin-Valat MJ, Vallet A, et al. In vivo phage display to identify new human antibody fragments homing to atherosclerotic endothelial and subendothelial tissues[J]. Am J Pathol, 2012, 180:2576-2589. |
| [19] | Chen Z, Wei J, Hao L, et al. Discovery of peptide ligands for hepatic stellate cells using phage display[J]. Mol Pharm, 2015, 12:2180-2188. |
| [20] | Huang XL, Wu J, Su TT, et al. Long-circulating liposomal daptomycin enhances protection against systemic methicillinresistant Staphylococcus aureus infection with improved therapeutic potential[J]. Acta Pharm Sin (药学学报), 2014, 49:701-710. |
| [21] | Strauss J, Kadilak A, Cronin C, et al. Binding, inactivation, and adhesion forces between antimicrobial peptide cecropin P1 and pathogenic E.coli[J]. Colloids Surf B, 2010, 75:156-164. |
| [22] | Sabiiti W, May RC, Pursall ER. Experimental models of cryptococcosis[J]. Int J Microbiol, 2012, 2012:626745. |
| [23] | Stie JL, Fox D. Blood-brain barrier invasion by Cryptococcus neoformans is enhanced by functional interactions with plasmin[J]. Microbiology, 2012, 158:240-258. |
| [24] | Tseng HK, Huang SH, Shackleford G, et al. The bloodbrain barrier:targeting in the pathogenesis of C. neoformans meningitis and drug discovery[J]. Anti-Infect Agents, 2013, 11:1-10. |
| [25] | Chen SH, Stins MF, Huang SH, et al. Cryptococcus neoformans induces alterations in the cytoskeleton of human brain microvascular endothelial cells[J]. J Med Microbiol, 2003, 52:961-970. |
| [26] | Shao K, Yu Z, Ning D, et al. Functionalized nanoscale micelles with brain targeting ability and intercellular microenvironment biosensitivity for anti-intracranial infection applications[J]. Adv Health Mater, 2015, 4:291-300. |
| [27] | Goyal AK, Garg T, Rath G, et al. Development and characterization of nanoembedded microparticles for pulmonary delivery of antitubercular drugs against experimental tuberculosis[J]. Mol Pharm, 2015, 12:3839-3850. |