 2016, Vol. 51
2016, Vol. 51



2. 复旦大学药学院 生药学教研室, 上海 201203
2. Department of Pharmacognosy, School of Pharmacy, Fudan University, Shanghai 201203, China
狼疮(lupus) 是一种慢性自身免疫性疾病,其临床表现多样,可从疲劳、轻微皮疹到肾衰竭。狼疮的发病机制十分复杂,受基因、环境和激素多方面影响,至今仍然不能明确[1]。辅助性T细胞(helper T cells,Th) 各亚群中,Th1介导干扰素-γ (IFN-γ) 的分泌,Th2和调节性T细胞 (regulatory T cell,Treg) 介导白细胞介素-10 (IL-10) 的分泌,Th17介导IL-17的分泌。Th各亚群通过分泌相关细胞因子,参与狼疮的发生发展[2, 3]。目前临床上治疗狼疮的药物主要为皮质类固醇激素、抗疟药物和细胞毒性药物。虽然这些药物改善和提高了狼疮的预后,但是疗效不理想,并且带有明显的细胞毒作用,包括血细胞数减少、感染和恶性肿瘤患病概率的增加[4]。2011年,美国食品药品监督管理局 (FDA) 批准贝利单抗治疗狼疮,该药作为人单克隆抗体,通过中和B淋巴细胞刺激因子产生疗效,但仍 对很大一部分患者的治疗效果不佳[5]。
柴胡是我国的传统中药和常用中药,其性味苦凉,有疏散退热、疏肝解郁、升阳举气之功效,用于治疗寒热往来、胸满胁痛、口苦耳聋和头痛目眩等[6]。以柴胡为主药的小柴胡汤性较平和,具有和解少阳之功,以清泄枢机之热为特点,有报道用于热毒炽盛型狼疮,疗效显著[7]。本课题组前期研究表明[8, 9, 10, 11],柴胡多糖离体具有抑制补体过度激活,调节巨噬细胞分泌和吞噬功能。狼疮小鼠灌胃给予柴胡多糖可明显改善肾病理损伤,减少肾免疫复合物沉积,降低尿蛋白量,降低小鼠体内多种抗核抗体水平[8, 12]。
目前,国内外抗狼疮药物研究中采用的动物模型有自发型模型 (如MRL/lpr)[13]和空肠弯曲菌 (Campylobacter jejuni-S131,CJ-S131) 诱导型模型[8]等。但是动物模型的实验周期均较长,不能实现药物离体筛选。
本研究通过CJ-S131离体再刺激狼疮样模型小鼠脾细胞,建立离体狼疮样小鼠炎症模型; 通过离体给予柴胡多糖,验证该模型是否能用于药物筛选和机制研究,并进一步观察柴胡多糖是否通过调控狼疮相关细胞因子的分泌,降低抗自身抗体水平来发挥药效。
材料与方法 实验动物健康SPF级BALB/c小鼠,雄性,18~20 g,由上海西普尔-必凯实验动物有限公司提供 (许可证号为: SCXK2008-0016)。所有实验方案均由复旦大学药学院动物伦理协会批准 (药论审第2011-7号)。
药品与试剂 药物的配制小叶黑柴胡多糖 (Bupleurum smithii var. parvifolium polysaccharides,BPs),由复旦大学药学院生药教研室提供,采用水提醇沉法从干燥的柴胡根块中提取柴胡多糖,并测得其提取率为3.2%,且为褐色粉末,溶于水。以D-半乳糖为标准品,采用硫酸-苯酚法测得柴胡粗多糖中总多糖含量为74.8%; 以D-半乳糖醛酸为标准品,经间羟基联苯显色测得柴胡粗多糖中糖醛酸含量为41.5%[8]。精密称取小叶黑柴胡多糖灭菌,加RPMI 1640培养液配成柴胡多糖800 μg·mL-1,分装,-20 ℃保存。以前期实验室工作为参考[14],培养体系中BPs终质量浓度分别设为5、10、20和40 μg·mL-1。
佐剂的配制参考文献[15]所述方法配制佐剂。称取羊毛脂10 g,石蜡油40 mL,融化混匀,150 ℃干热灭菌1 h,配制不完全弗氏佐剂 (Freund’s incomplete adjuvant,IFCA)。用IFCA加入200 mg·mL-1 Bacilue Calmette-Guérin,配制10 g·L-1弗氏完全佐剂 (Freund’s complete adjuvant,FCA)。
含CJ-S131的弗氏完全佐剂空肠弯曲菌灭活菌株 (CJ-S131),购自上海市卫生防疫站 (批号2011-11-2)。将甲醛化空肠弯曲菌CJ-S131株菌悬液用无菌生理盐水洗3次,在无菌条件下调节菌悬液浓度为3.6×109 CFU·L-1,80 ℃灭菌30 min,-20 ℃保存。取已灭菌的菌悬液,用RPMI1640稀释至3.6×106 CFU·L-1,每管1 mL分装,-20 ℃保存,用于细胞培养。
试剂RPMI 1640培养液和胎牛血清 (FBS) (Gibco公司); MTT (Sigma公司); IFN-γ、IL-10 ELISA试剂盒 (船夫公司); IL-17 ELISA试剂盒 (NeoBioscience Technology Co.,Ltd.); 羊抗小鼠IgG干粉 (创赛生物工程有限公司); 羊抗小鼠IgG-HRP (长岛生物技术公司); dsDNA (小牛胸腺DNA,Sigma公司),其他试剂为国产分析纯。
模型制备及分组根据文献[15]建立模型。小鼠随机分为3组,每组4只动物: 正常对照组 (Nil)、佐剂对照组 (FCA) 和模型组(model)。
初次免疫: Day 0,正常对照组小鼠于右足皮下注射生理盐水 (NS) 50 μL; 佐剂对照组注射不含CJ-S131仅含NS的FCA 50 μL; 模型组注射免疫用含CJ-S131佐剂 (即FCA+CJ-S131) 50 μL。第二次免疫: Day 14,正常对照组和佐剂对照组每只小鼠尾静脉注射NS 0.2 mL; 模型组每只小鼠尾静脉注射CJ-S131细菌悬液0.2 mL。Day 19处死小鼠取脾细胞进行后续实验。
柴胡多糖对狼疮样小鼠脾脏细胞分泌功能的影响脱颈处死小鼠,释放脾细胞,调节细胞数至每毫升8×106个加入48孔板中,每孔250 μL细胞溶液和250 μL药物 [含CJ-S131培养基500 μL (CJ-S131终浓度为1×105 CFU·mL-1) 或培养基],每孔总体积1 000 μL。各孔中按以下分组情况加样: ① 不加刺激剂和药物的对照组: 只加RPMI 1640 + 10% FBS培养基; ② CJ-S131刺激组: 加入含CJ-S131培养基; ③ BPs与CJ-S131联合处理组: 每孔加含药物和CJ-S131的培养基 (BPs终质量浓度为5、10、20和40 μg·mL-1); ④ 药物BPs单独处理组: BPs终质量浓度为5、10、20和40 μg·mL-1。每组3个复孔,37 ℃、5% CO2中培养 48 h后离心,吸取每孔的细胞上清液,分装后放入-80 ℃冰箱保存,待测。
细胞培养上清中总IgG的检测[16, 17]10 μg·mL-1羊抗小鼠IgG ,以每孔100 μL加入96孔酶标板,4 ℃过夜。第2天,倾去上清,PBS-1% BSA封闭 (每孔100 μL), 37 ℃孵育2 h; 分别加入不同浓度的纯小鼠IgG ,或待测1∶5稀释的细胞培养上清 (每孔100 μL),37 ℃孵育2 h,加洗涤液250 μL,洗涤5次; 加1∶5 000稀释的羊抗小鼠IgG-HRP酶标抗体100 μL,37 ℃孵育2 h; 清洗后加入邻苯二胺 (OPD) 底物溶液,37 ℃避光孵育20 min; 每孔加入50 μL 2 mol·L-1 H2SO4溶液终止反应,用酶标仪于492 nm处测吸光度值。同时以纯小鼠IgG作标准曲线,计算小鼠总IgG量。
细胞培养上清中抗双链DNA (dsDNA) 抗体的检测[16, 17]dsDNA的测定步骤同上。50 μg·mL-1 dsDNA 100 μL加入酶标板。第2天封闭后加待测细胞培养上清原液,加1∶1 000稀释的羊抗小鼠IgG-HRP。用酶标仪于492 nm处测定吸光度值。
细胞培养上清中狼疮相关细胞因子的检测[16, 17]采用ELISA试剂盒,检测培养上清中IFN-γ、IL-10和IL-17的水平,测定IFN-γ和IL-10时,细胞培养上清以1∶2稀释; 测定IL-17时,细胞培养上清用原液测定。具体步骤按试剂盒说明书进行,450 nm测定吸光度值,建立各个细胞因子的标准曲线,并计算各实验组细胞培养上清IFN-γ、IL-10和IL-17的水平。
柴胡多糖对狼疮样小鼠脾脏细胞增殖活性的影响按照“柴胡多糖对狼疮样小鼠脾脏细胞分泌功能的影响”的方法分组和培养细胞,每组4个复孔,37 ℃、5% CO2培养箱培养48 h,每孔加5 g·L-1 MTT 50 μL,继续培养4 h。每孔加MTT溶解液500 μL,37 ℃保温振荡过夜,第2天于570 nm处测定吸光度值。
统计学分析
实验数据以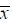 ± s表示,采用Statview软件进行单因素方差分析比较组间差异性,P < 0.05为具有显著性差异。
± s表示,采用Statview软件进行单因素方差分析比较组间差异性,P < 0.05为具有显著性差异。
未加CJ-S131再刺激时,模型组培养上清中总IgG的水平均已显著高于正常对照组和佐剂对照组 (P < 0.001); 而给予CJ-S131再次刺激脾细胞后,3组的总IgG水平均较未刺激时显著升高,而模型组的总IgG水平升高幅度更大,且与两对照组相比均有显著性差异 (P < 0.001) (图 1A)。

|
Figure 1 Effect of Bupleurum smithii var. parvifolium polysaccharides (BPs) on total IgG production in spleen of mice. The mice were randomly divided into three groups: the Nil group received normal saline; the FCA gruop was immunized with Freund’s complete adjuvant; the model group was immunized with emulsifier containing Freund’s complete adjuvant and Campylobacter jejuni strain (CJ-S131). Mice were sacrificed on Day 19 and the splenocytes were treated with BPs at different concentrations and stimulated with or without CJ-S131. The culture supernatant of splenocytes was collected and diluted at 1∶5 for total IgG detection. Without BPs treatment (A),and with BPs treatment in groups of Nil (B),FCA (C) and Model (D). n = 3,± s. ###P < 0.001 vs Nil group; ΔΔΔP < 0.001 vs FCA group; P < 0.05,**P < 0.01,***P < 0.001 vs no BPs added group
|
未加CJ-S131刺激,单独加入BPs,能轻度抑制正常对照组 (图 1B) 和佐剂对照组 (图 1C) 的总IgG水平(P < 0.05),显著地抑制模型组 (图 1D) IgG水平 (P < 0.001)。
加入CJ-S131刺激后,各组IgG水平均较未刺激前显著升高,且BPs均能显著抑制各组高水平IgG (P < 0.001) (图 1B~D)。
2 柴胡多糖对狼疮样小鼠脾细胞分泌抗dsDNA抗体水平的影响未加CJ-S131刺激时,模型组与佐剂对照组相比,培养基中已有较高抗dsDNA抗体水平 (P < 0.01); 而加入CJ-S131再刺激脾细胞后,模型组的抗dsDNA抗体水平进一步升高,且与对照组相比均有显著性差异 (P < 0.01)。CJ-S131对两对照组dsDNA水平无明显升高作用 (图 2A)。

|
Figure 2 Effect of BPs on anti-dsDNA production in spleen of mice. The culture supernatant of splenocytes was collected for anti-dsDNA antibody detection. Without BPs treatment (A),and with BPs treatment in groups of Nil (B),FCA (C) and Model (D). n = 3,± s. ##P < 0.01 vs Nil group; ΔΔΔP < 0.001 vs FCA group; P < 0.05,**P < 0.01 vs no BPs adding in CJ-S131 stimulating group
|
未加CJ-S131刺激,单独加入BPs对各组小鼠脾细胞的dsDNA水平无明显影响 (图 2B~D)。
CJ-S131刺激升高模型组细胞的抗dsDNA抗体的分泌水平,且20~40 μg·mL-1 BPs明显抑制该升高作用 (P < 0.05) (图 2A,D)。
3 柴胡多糖对狼疮样小鼠脾细胞分泌IFN-γ的影响未加CJ-S131刺激时,细胞培养上清的IFN-γ水 平在模型组和对照组之间无差异; 加入CJ-S131再刺激脾细胞后,各组的IFN-γ水平较刺激前均显著升 高,且模型组的IFN-γ升高幅度最大,约为刺激前的50倍 (P < 0.001) (图 3A)。

|
Figure 3 Effect of BPs on IFN-γ production in spleen of mice. The culture supernatant of splenocytes was collected and diluted at 1:2 for IFN-γ detection. Without BPs treatment (A),and with BPs treatment in groups of Nil (B),FCA (C) and Model (D). n = 3,± s. ###P < 0.001 vs Nil group; ΔΔΔP < 0.001 vs FCA group; ***P
< 0.001 vs no BPs adding in CJ-S131 stimulating group
|
虽然单独加入BPs对各组的IFN-γ分泌没有明 显影响,但BPs能显著抑制CJ-S131刺激引发的各组IFN-γ水平的升高(图 3B~D)。
4 柴胡多糖对狼疮样小鼠脾细胞分泌IL-10的影响未加CJ-S131刺激时,IL-10的水平在模型组和对照组之间无差异; 而离体CJ-S131再刺激脾细胞后,各组IL-10的水平较刺激前均显著升高,且模型组的IL-10升高幅度最大,约为刺激前的70倍 (P < 0.001) (图 4A)。

|
Figure 4 Effect of BPs on IL-10 production in spleen of mice. The culture supernatant of splenocytes was collected and diluted at 1:2 for IL-10 detection. Without BPs treatment (A),and with BPs treatment in groups of Nil (B),FCA (C) and Model (D). n = 3,± s. ###P < 0.001 vs Nil group; ΔΔΔP < 0.001 vs FCA group; ***P < 0.001 vs no BPs adding in CJ-S131 stimulating group
|
BPs单独加入对各组的IL-10分泌没有影响。BPs能显著抑制CJ-S131加入后各组IL-10水平的升高 (P < 0.001) (图 4B~D)。
5 柴胡多糖对狼疮样小鼠脾细胞分泌IL-17的影响未加CJ-S131刺激时,IL-17水平在模型组和对照组之间无显著性差异; 加入空肠弯曲菌CJ-S131再刺 激后,正常对照组和佐剂对照组的IL-17水平无明显改变,模型组的IL-17水平较对照组均显著升高 (P < 0.001),约为两对照组的6倍 (表 1)。但是同时给予BPs,各浓度均对IL-17的升高无明显抑制作用 (数据未在文章中列出)。
|
|
Table 1 IL-17 production and proliferation of splenocytes of lupus like syndrome mice. The culture supernatant of splenocytes was collected for IL-17 detection; splenocytes proliferation were determinted by MTT assay. n = 3 (IL-17),n = 4 (proliferation),± s. ##P < 0.01,###P < 0.001 vs Nil group; ΔΔP < 0.01,ΔΔΔP < 0.001 vs FCA group; ***P < 0.001 vs without CJ-S131 group
|
未加CJ-S131刺激时,模型组与正常对照组、佐剂对照组相比,其细胞活性均显著升高 (P < 0.01); 而加入CJ-S131再刺激脾细胞后,正常对照组、佐剂对照组和模型组的细胞活性均较再刺激之前显著升高 (P < 0.001),且模型组活性与佐剂对照组相比具有显著性差异 (P < 0.01) (表 1)。柴胡多糖各剂量组对脾细胞活性均无显著影响 (数据未在文章中列出)。
讨论狼疮是一种难治、反复发作、致死率高的严重自身免疫病[18]。虽然狼疮的病因和发病机制尚未完全阐明,但患者均存在免疫调节功能的严重失调,机体高水平的抗自身抗体和低水平的单核巨噬细胞清除能力,使免疫复合物增多,并广泛性激活补体系统,造成机体的炎症扩大和组织器官损伤,同时使机体对自身抗原的识别长期存在,造成病情的迁延不愈[19]。
狼疮患者可查到多种自身抗体的产生,如抗核抗体、抗dsDNA抗体和抗组蛋白抗体等,高抗自身抗体是机体组织受到攻击,发生损伤的重要原因。机体免疫功能异常,细胞因子分泌的改变,如IFN-γ、IL-10和IL-17等水平升高[18, 20],参与打破免疫耐受,形成免疫系统对自身的攻击,机体产生大量的抗自身抗体,维持了机体的高炎症水平,最终造成了组织病理损伤。
由于空肠弯曲菌CJ-S131的43 kDa的热休克蛋白和脂多糖均具有与机体自身抗原相似的抗原表位 (分子模拟现象),细菌成分作为交叉抗原进入体内,产生应激,引起一系列免疫反应,T细胞活化后通过旁路途径辅助B细胞多克隆激活,引起自身抗体的产生,使得CJ-S131打破小鼠的免疫耐受状态,诱导小鼠形成具有典型的狼疮样症状,是研究狼疮病理机制及寻找治疗药物较理想的模型之一[21, 22]。
通过两次给予小鼠CJ-S131免疫,可以打破小鼠的免疫耐受,较短时间内诱导小鼠形成狼疮样高抗自身抗体,建立模型。脾为机体重要的外周免疫器官,是T细胞和B细胞定居的场所,还有一定量的单核-巨噬细胞。CJ-S131两次刺激小鼠,在脾脏可形成针对CJ-S131的免疫应答细胞。本研究通过取脾细胞,离体再次给予CJ-S131的刺激,可建立离体狼疮炎症模型。
与正常对照组和佐剂对照组相比,未加CJ-S131的模型组脾细胞活跃度较高; 在培养上清中,模型组有较高的抗dsDNA抗体和总IgG水平; 但细胞因子水平各组间差异不大。而各组离体脾细胞经CJ-S131再刺激后,脾细胞活跃度明显增高,总IgG水平和细胞因子水平均升高,但以模型组增高更加显著。
狼疮患者多克隆B细胞过度活化,促使B细胞分泌大量免疫球蛋白,致使患者体内免疫球蛋白升高,其中以总IgG水平升高最为明显[23]。在本实验中,无论是否加入CJ-S131再刺激,柴胡多糖各剂量组 (5~40 μg·mL-1) 对两对照组和模型组的总IgG水平均有明显抑制作用。
抗dsDNA抗体是狼疮患者体内的特异性自身抗体,几乎只出现在狼疮中,在狼疮的发生发展中起着重要的作用[24]。抗dsDNA可与相应抗原结合形成抗原抗体免疫复合物,后者可激活补体系统[25]。研究发现,BPs同前期整体实验结果类似[8],BPs (20和40 μg·mL-1) 能够一定程度降低CJ-S131刺激产生的高水平抗dsDNA抗体。
IFN-γ在免疫应答初始阶段主要由自然杀伤细胞分泌,在适应性免疫中主要由Th1细胞分泌。研究表明,在狼疮患者血清中可检测到IFN-γ水平的上升[20],且该水平与疾病活动性呈正相关,因此IFN-γ是一种典型的Th1型细胞因子,它可以提高细胞免疫应答水平,增强B细胞活化从而加重狼疮肾炎[26]。离体再刺激之前,IFN-γ的水平在模型组和对照组之间无差异; 而离体CJ-S131再刺激脾细胞后,正常组、佐剂对照组和模型组的IFN-γ的水平较刺激前均显著升高,且模型组的IFN-γ升高幅度最大。BPs能显著抑制CJ-S131刺激导致的各组IFN-γ水平升高,最高抑制率达97%。
Treg和Th2细胞都参与IL-10的分泌,IL-10可以刺激B细胞,促进自身抗体的合成,因此在很多狼疮患者的血浆中IL-10的水平与疾病活动和抗dsDNA水平呈正相关[3]。CJ-S131再刺激之前,IL-10的水平在模型组和对照组之间无明显差异; 而离体CJ-S131再刺激脾细胞后,各组的IL-10的水平较刺激前均显著升高,且模型组的IL-10升高幅度最大。BPs能显著抑制CJ-S131刺激后各组升高的IL-10水平,最高抑制率达90%。
IL-17主要由Th17细胞产生,是一种重要的前炎性细胞因子,能够单独或与B细胞活化因子协同促进B细胞增殖、存活及自身抗体的产生[27]。在皮肤损害和神经系统损害的狼疮患者中,尤其是狼疮肾炎患者体内可测到高水平的IL-17[28]。本研究结果表明,离体再刺激之前,IL-17的水平在模型组和对照组之间无显著性差异; 离体CJ-S131再刺激后,模型组的IL-17的水平较对照组均显著升高。而BPs各浓 度对模型组IL-17的升高水平均无显著影响,这说明BPs治疗狼疮的作用靶点可能不在Th17细胞。
BPs各剂量对狼疮样小鼠脾脏细胞的增殖活性没有影响,表明BPs对细胞没有明显的毒副作用,主要不是通过细胞毒性来发挥作用的。
补体系统过度激活、单核/巨噬细胞功能异常及淋巴细胞的调节功能异常皆可导致或加剧狼疮。前期研究已经观察到,BPs具有抑制补体系统过度激活和调节单核/巨噬细胞功能的作用[8, 9, 10]。本项研究表明,BPs对淋巴细胞功能异常也具有调节作用,对T细胞亚群中Th1、Th2和Treg的分泌功能,均具有调节作用,但对Th17亚群分泌的IL-17没有明显影响。
通过本研究,建立了具有狼疮样病变的离体脾细胞炎症模型,在该模型上进一步观察了BPs防治狼疮作用的靶点,为进一步药物开发提供了思路和研究平台。
| [1] | Vilas-Boas A, Morais SA, Isenberg DA. Belimumab in systemic lupus erythematosus[J]. RMD Open, 2015, 1:e000011. |
| [2] | Talaat RM, Mohamed SF, Bassyouni IH, et al. Th1/Th2/Th17/Treg cytokine imbalance in systemic lupus erythematosus (SLE) patients:correlation with disease activity[J]. Cytokine, 2015, 72:146-153. |
| [3] | Robinson ES, Werth VP. The role of cytokines in the pathogenesis of cutaneous lupus erythematosus[J]. Cytokine, 2015, 73:326-334. |
| [4] | Yu YY, La Cava A. Current drugs in systemic lupus erythematosus[J]. Drug Develop Res, 2011, 72:561-572. |
| [5] | Stohl W, Hilbert DM. The discovery and development of belimumab:the anti-BLyS-lupus connection[J]. Nat Biotechnol, 2012, 30:69-77. |
| [6] | Li F, Li JB, Zhang DM. Pharmacological research progress on Bupleurum[J]. Lishizhen Med Mater Med Res (时珍国医国药), 2004, 15:120-121. |
| [7] | Wang BJ, Wang YN, Wang QR, et al. Western medicine and TCM heat pattern analysis of 181 cases of systemic lupus erythematosus[J]. Neimonggu J Tradit Chin Med (内蒙古中医药), 2003, S1:27. |
| [8] | Wang Z, Li H, Xu H, et al. Beneficial effect of Bupleurum polysaccharides on autoimmune disease induced by Campylobacter jejuni in BALB/c mice[J]. J Ethnopharmacol, 2009, 124:481-487. |
| [9] | Wang Z, Cheng XQ, Xu H, et al. Effects of Bupleurum smithii var. parvifolium total polysaccharides on immune functions in mice[J]. Chin J Clin Pharm (中国临床药学杂志), 2009, 18:200-204. |
| [10] | Wu L, Jiang L, Zhang YY, et al. Effects of polysaccharides from Bupleurum smithii var. parvifolium on the functions of murine peritoneal macrophages[J]. Chin J Clin Pharm (中国临床药学杂志), 2013, 22:1-5. |
| [11] | Xu H, Zhang Y, Zhang J, et al. Isolation and characterization of an anti-complementary polysaccharide D3-S1 from the roots of Bupleurum smithii[J]. Int Immunopharmacol, 2007, 7:175-182. |
| [12] | Jiang YW, Li H, Zhang YY, et al. Beneficial effect of Bupleurum polysaccharides on autoimmune-prone MRL-lpr mice[J]. Clin Dev Immunol, 2012, 2012:842928. |
| [13] | Swenson J, Olgun S, Radjavi A, et al. Clinical efficacy of buprenorphine to minimize distress in MRL/lpr mice[J]. Eur J Pharmacol, 2007, 567:67-76. |
| [14] | Wu J, Zhang YY, Guo L, et al. Bupleurum polysaccharides attenuates lipopolysaccharide-induced inflammation via modulating Toll-like receptor 4 signaling[J]. PLoS One, 2013, 8:e78051. |
| [15] | Lu Y, Ou YY, Li H. Optimization of a mouse model of systemic lupus erythematosus induced by Campylobacter jejuni-S131[J]. Chin J Clin Pharm (中国临床药学杂志), 2014, 23:343-348. |
| [16] | Li T, Wang H, He S. Induction of interleukin-6 release from monocytes by serine proteinases and its potential mechanisms[J]. Scand J Immunol, 2006, 64:10-16. |
| [17] | Wang JB, Zhou X, Wang ZW, et al. Effect of tetanus toxoid powder needleless injection on the concentration of serum antibody IgG in mice[J]. Acta Pharm Sin (药学学报), 2009, 44:1406-1409. |
| [18] | Choi J, Kim ST, Craft J. The pathogenesis of systemic lupus erythematosus-an update[J]. Curr Opin Immunol, 2012, 24:651-657. |
| [19] | Kavai M, Szegedi G. Immune complex clearance by monocytes and macrophages in systemic lupus erythematosus[J]. Autoimmun Rev, 2007, 6:497-502. |
| [20] | Davis LS, Hutcheson J, Mohan C. The role of cytokines in the pathogenesis and treatment of systemic lupus erythematosus[J]. J Interferon Cytokine Res, 2011, 31:781-789. |
| [21] | Zang XX, Ma BL, Bai J, et al. HSP43 of Campylobacter jejuni induced autoantibodies production[J]. Chin J Immunol (中国免疫学杂志), 1996, 12:279-282. |
| [22] | Sun B, Ma BL. Research on the efficacy mechanism of TWP on autoimmunity disease induced by Campylobacter jejuni[J]. Chin Pharm J (中国药学杂志), 1993, 28:724-727. |
| [23] | Zhao XD. Change of T-lumphocyte subsets, immunoglobulin and complement level in peripheral blood of patients with systemic lupus erythematosus[J]. Guide China Med (中国医药指南), 2010, 8:25-26. |
| [24] | Chen XG, Zhang XL, Liu CY, et al. Clinial significance of changes of expression of anti-dsDNA antibody in serum in patients with SLE[J]. J Radioimmunol (放射免疫学杂志), 2008, 21:6-8. |
| [25] | Guo XB, Wang P, Liu CH. Detection of immunoglobulins and C3 in SLE patients[J]. J Henan Med Coll Staff Work (河南职工医学院学报), 2007, 19:481-482. |
| [26] | Zeng HQ, Ye ZZ, Wang X, et al. Correlation between the level of IFN-γ in peripheral blood of patients with SLE and the activity of disease[J]. Curr Immunol (现代免疫学), 2007, 27:69-72. |
| [27] | Huang ZF, Li XL, Li HX, et al. The role of Th17 cells and interleukin 17 in the pathogenesis of systemic lupus erythematosus[J]. Med J Air Force (空军医学杂志), 2014, 30:28-30. |
| [28] | Li D, Guo B, Wu H, et al. Interleukin-17 in systemic lupus erythematosus:a comprehensive review[J]. Autoimmunity, 2015, 48:353-361. |