



2016年全国农村可再生能源统计表明, 我国农作物秸秆的秸秆综合利用率达到81.68%[1]。秸秆的利用途径为肥料化、饲料化、燃料化、基料化和原料化, 利用率分别为47.20%、17.99%、11.79%、2.23%、2.47%[2]。中国产业信息网发布的2018年全国水稻产量为2.027×108 t, 根据毕于运提出的我国水稻草谷比平均值为0.90[3], 可以推算2018年我国全年水稻秸秆产量为1.824×108 t。稻秸是我国主要的作物秸秆类型, 我国水稻秸秆年产量大, 探索利用稻秸的新途径, 减轻对环境的压力, 是急需解决的难题。
Martinez等[4]报道与饲喂相同饲料的瘤胃液供体羊相比, 体外Rusitec发酵培养16 d后, 自动核糖体基因间隔分析(ARISA)结果表明, 未检测到特异性细菌群落的变化; 而Zapletalova等[5]证明了利用双外流连续培养技术, 在接种瘤胃内容物10 d内, 瘤胃细菌群落结构发生了变化, 特别是提高了厚壁菌门(Firmicutes):拟杆菌门(Bacteroidetes)的比例。华金玲等[6]以尼龙袋试验表明, 水稻秸秆的72 h瘤胃降解率为35.41%;张轶凤等[7]以山羊瘤胃液发酵稻秸, 72 h尼龙袋瘤胃降解率为30.57%。因此, 根据前人的研究表明, 稻秸的体外72 h瘤胃降解率较低。而Lin等[8]以柳枝稷和DDGS为主要碳氮源, 对瘤胃液进行长时间的频繁接种, 结果表明即使在低消化率生物质为主要底物的情况下, 来自瘤胃的混合培养细菌群落依然可以在体外维持。瘤胃液和厌氧污泥共同接种稻秸培养196 d的结果也表明, 稻秸的木质纤维素在体外被成功高效地降解[9]。
目前厌氧发酵稻秸成为生产SCFA是的一种新型的利用方向, 在替代以石化为原料的不可再生资源中作为化学平台原料具有优势。己酸是瘤胃发酵纤维素产生的SCFA之一, 己酸在抗菌剂、饲料添加剂、风味剂、化工原料方面有较大的应用潜力[10-13], 且经济价值是乙醇的10倍以上。Lin等[8]报道体外发酵柳枝稷时添加乙醇可以显著提高乙酸和己酸的产量, 在有机废弃物中添加乙醇可以显著提高己酸的产量[14], 说明乙醇是己酸合成的主要供氢体[15]。目前, 长期培养和连续接种对稻秸厌氧发酵生产SCFA的产量和微生物群落的影响少有报道。
本文研究了水稻秸秆与瘤胃液共培养8代后SCFA的产量变化, 同时还研究了添加乙醇对瘤胃细菌发酵稻秸生产SCFA的变化, 并进一步阐述了细菌群落结构特征, 以期为揭示瘤胃细菌发酵稻秸产酸的特点, 并为稻秸的利用提供参考依据。
1 材料与方法 1.1 材料水稻秸秆和玉米酒精糟(DDGS)来自扬州大学动物营养与饲料工程技术研究中心。稻秸经65℃烘干48 h后得到风干样品, 再粉碎过0.50 mm筛, 密封备用。体外培养底物以0.80 g稻秸和0.16 g DDGS配制而成。瘤胃液来自扬州大学实验农牧场(高邮)的带有永久性瘤胃瘘管的山羊。在晨饲前采集瘤胃内容物, 经4层无菌纱布过滤后, 迅速带回实验室。
培养缓冲液参照Menke(1979)[16]配制:微量元素溶液(A液, 1 L):13.2 g CaCl2·2H2O, 10.0 g MnCl2·4H2O, 1.0 g CoCl2·6H2O, 8.0 g FeCl3·6H2O;缓冲溶液(B液, 1 L):4.0 g NH4HCO3, 35.0 g NaHCO3;常量元素溶液(C液, 1 L):5.7 g Na2HPO4, 6.2 g KH2PO4, 0.6 g MgSO4·7H2O;0.1%(W/V)刃天青溶液。再以蒸馏水400 mL + A液0.1 mL + B液200 mL + C液200 mL +刃天青溶液1 mL的比例配制。现用现配。
试验使用150 mL体积的厌氧培养瓶为培养装置, 采集瘤胃液前称取稻秸和DDGS于培养瓶中, 加入15 mL缓冲液, 持续通入CO2 10 min, 加入2.5% (W/V)硫化钠溶液0.1 mL后, 胶塞密封, 光照至刃天青褪色。每瓶加入2 mL瘤胃液, 胶塞密封加铝盖后, 于39℃静置培养3 d, 视为第1代。以不加乙醇为对照组, 以加0.2 mL乙醇为试验组, 每组3个平行。
每3 d传代一次, 传代量为2 mL, 其它操作同上。共培养8代。
1.2 方法 1.2.1 样品采集与处理在每次传代前和3 d培养结束时, 以气压表(DPG1000B15PSIG-5, CeComp Electronics Inc.)测定瓶内气压(psi)。以pH计(PB- 21型, Sartorius)测定发酵液pH值。按Lin等[8]的方法对测定短链脂肪酸进行去蛋白操作:吸取0.6 mL发酵液, 加入含有0.3 mL硫酸铜溶液(12.5%, M/V, 含巴豆酸0.002%, M/V)和0.6 mL碳酸钙溶液(26.45%, M/V)的去蛋白溶液中, 漩涡混匀后在-20℃冰箱中冷冻过夜; 解冻后以12 000 r/min离心10 min, 经0.22 μm水相针式滤膜过滤, 4℃保存备测。收集第8代的发酵混合液, 按Lin等[8]方法分离细菌。具体方法为:用30 mL提取缓冲液(100 mmol/L tris/HCl, 10 mmol/L EDTA, 0.15 mol/L NaCl, pH 8.0)将样品(固液混合)转移至搅拌杯中, 高速搅拌2 min后完全无损经无菌2层纱布过滤入50 mL离心管, 在12 000 r/min离心30 min, 弃去上清液, 再用提取缓冲液复悬后, 送北京诺禾致源生物技术有限公司进行总细菌DNA的提取和高通量测序分析。
1.2.2 测定方法采用日本岛津GC-14B气相色谱法测定短链脂肪酸含量。气相色谱仪参数:色谱柱为毛细管柱, 柱温130℃, 汽化温度180℃, 氢离子火焰检测器, 检测温度180℃, 压力为60 KPa, 氢气压力为50 KPa, 氧气压力50 KPa, 灵敏度(档)为101, 衰减3.0, 以氮气为载气, 进样量1.0微升, 测定样品的乙酸、丙酸、异丁酸、丁酸、异戊酸、戊酸、己酸的含量。
细菌群落结构分析采用高通量测序Illumina Hiseq平台(北京诺禾致源生物技术有限公司)。引物选用16S V4区的515F和806R (515F 5'-GTGCCAGCMGCCGCGGTAA-3', 806R 5'-GGACTACHVGGGTWTCTAAT-3')进行扩增。按张雪姣[17]的方法处理序列, 使用Mothur软件以一致性(Identity)为97%将序列聚类成为OTUs, 绘制韦恩图分析共同存在的OTU和特有的OTU; 通过UPGMA(Unweighted pair-group method with arithmetic mean)绘制聚类树分析样本间的相似性; 选用LEfSe统计分析方法对分组样品的物种组成和群落结果进行差异显著性检验; 使用CCA(Canonical correspondence analysis)方法分析细菌群落与SCFA比例的相关性。
1.2.3 统计方法以SPSS 21.0软件中单因素ANOVA方法进行差异显著性分析, 以组别为变量, 用Tukey进行多重比较。对于不同处理指标, 在不服从正态分布时, 做非参数检验重新分析显著性。试验结果以"平均数±标准差"表示, 以P < 0.05表示差异显著, P < 0.01表示差异极显著。
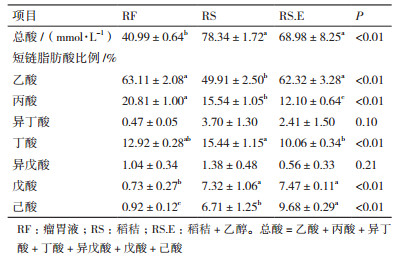
2 结果 2.1 乙醇对稻秸体外发酵的短链脂肪酸产量变化由表 1可见, 无论是瘤胃液还是体外发酵液, 乙酸是占比例最大的SCFA, 丙酸和丁酸的比例相近。经体外培养传代8次的稻秸发酵液的总短链脂肪酸产量显著高于瘤胃液(P < 0.05)。与未添加乙醇的稻秸发酵液相比, 添加乙醇显著提高了乙酸、戊酸和己酸的比例, 降低了丙酸和丁酸的比例(P < 0.01), 总SCFA产量及异丁酸和异戊酸比例无显著差异。
由图 1可知, 本试验共测定得到911个OTU, 共有的OTU个数为292, 占总OTU数量的32.05%。瘤胃液与稻秸发酵液共有的OTU为321个, 瘤胃液与添加乙醇的稻秸发酵液共有的OTU为322个, 而稻秸发酵液与添加乙醇的稻秸发酵液共有的OTU为549个。山羊瘤胃液共得到539个OTU, 稻秸发酵液得到639个OTU, 稻秸发酵液添加乙醇后共得到633 OTU。瘤胃液有188个特有的OTU, 占总OTU数量的20.64%, 稻秸发酵液有61个特有的OTU, 占总OTU数量的6.70%;添加乙醇的稻秸发酵液有54个特有的OTU, 占总OTU数量的5.92%。

|
| RF:瘤胃液;RS:稻秸;RS.E:稻秸+乙醇 图 1 稻秸体外发酵及添加乙醇的瘤胃细菌Venn图 |
使用UPGMA对样品进行基于Weighted Unifrac方法的门水平聚类分析, 结果见图 2。由图 2可知, 3个组可分为2大类, 其中瘤胃液和稻秸发酵液的细菌群落相似度较高, 添加乙醇的稻秸发酵液单独为一类, 说明体外传代8次的稻秸发酵液与瘤胃液亲缘关系较近, 而添加乙醇显著改变了细菌区系。进一步分析菌门的相对丰度可知, 山羊瘤胃液中以拟杆菌门(Bacteroidetes)和厚壁菌门(Firmicutes)为主要菌门(分别为72.35%和24.70%)。以稻秸和DDGS为发酵底物, 连续传代8次后, 发酵液的拟杆菌门相对丰度下降, 厚壁菌门相对丰度升高(分别为57.98%和32.71%), 且添加0.2mL乙醇显著提高了厚壁菌门和放线菌门(Actinobacteria)的相对丰度(分别为51.91%和12.27%, P < 0.01)。

|
| RF:瘤胃液;RS:稻秸;RS.E:稻秸+乙醇 图 2 稻秸体外发酵及添加乙醇的瘤胃细菌群落间相似性聚类树 |
本试验选取相对丰度前20的细菌, 进行属水平相对丰度的比较(图 3)。由图 3可见, 瘤胃液中有76.24%的细菌无法用数据库定性。瘤胃液中未定性的理研菌属(unidentified Rikenellaceae)和未定性的普雷沃氏菌属(unidentified Prevotellaceae)相对丰度分别为0.64%和6.35%, 体外培养使这两个菌属的相对丰度显著升高(39.82%和13.10%, P < 0.05), 但添加乙醇使该菌属相对丰度显著降低(0.42%和1.84%, P < 0.05)。与瘤胃液相比, 双歧杆菌属(Bifidobacterium)的相对丰度在秸秆发酵液中无显著变化(0.17%和1.65%), 但添加乙醇显著提高了该菌属的相对丰度(24.82%, P < 0.01)。同样地, 添加乙醇显著提高了未定性的毛螺菌属(unidentified Lachnospiraceae)、产琥珀酸菌属(Succiniclasticum)、脱硫弧菌菌属(Desulfovibrio)和未定性的梭菌属(unidentified Clostridiales)的相对丰度(P < 0.05)。相反, 秸秆与瘤胃液体外共培养使未定性的瘤胃球菌属(unidentified Ruminococcaceae)和未定性的拟杆菌属(unidentified Bacteroidales)相对丰度降低(P < 0.01)。

|
| RF:瘤胃液;RS:稻秸;RS.E:稻秸+乙醇 图 3 乙醇对瘤胃细菌相对丰度的影响(属水平,相对丰度前20) |
稻秸发酵液的LEfSe分析柱状图和进化分支图见图 4, LDA score(柱形长度)大小代表差异物种的影响大小。由图可知, 添加乙醇后出现了不同的Biomarker, 且乙醇使显著性差异物种数增加。在门水平上, 稻秸发酵液中仅拟杆菌门内的细菌存在显著差异; 但添加乙醇后, 3个主要门的细菌存在显著差异, 分别为厚壁菌门、放线菌门和变形菌门。进一步分析可得, 稻秸发酵液中有9个特异性的Biomarker, 根据LDA score(柱形长度)的大小, 依次为拟杆菌(纲、目、属)、理研菌(科、属)、普雷沃氏菌(科、属)、栖瘤胃普雷沃氏菌和拟杆菌RM68。而添加乙醇后有21个特异性的Biomarker, 根据LDA score(柱形长度)的大小, 依次为双歧杆菌(纲、科、属)、放线菌(纲、属)、厚壁菌属、梭菌芽孢杆菌(纲、目)、毛螺旋菌科、脱硫弧菌(纲、目、属)、变形菌(门、纲)、丹毒丝菌(纲、目、科)、Solobacterium属、氢化厌氧细菌属、硫酸盐还原菌和毛螺旋菌AC2012。

|
| RF:瘤胃液;RS:稻秸;RS.E:稻秸+乙醇 图 4 稻秸体外发酵及添加乙醇的瘤胃细菌群落间差异LEfSe分析图(A)和进化分支图(B) |
为分析发酵液细菌群落与发酵参数间的关系, 以细菌群落在门水平上的相对丰度数据为物种数据, 发酵液乙酸、丙酸、丁酸、戊酸和己酸比例为变量, 进行了CCA分析(图 5)。由图 5可知, 稻秸发酵液的3个平行样本(RS1、RS2、RS3)较集中, 添加乙醇的稻秸发酵液的3个平行样本(RS.E1、RS.E2、RS.E3)也较集中, 说明组内3个样本的群落结构相近, 但RS和RS.E相距较远, 说明乙醇对细菌群落结构的影响较大。第1排序轴和第2排序轴分别对稻秸发酵液的细菌群落变化解释率为51.46%和41.9%, 前两轴共解释了总变异的93.36%, 在对细菌群落的解释中起主导作用。其中丙酸比例(r=- 0.67)、戊酸比例(r=0.82)和己酸比例(r=0.80)与第1排序轴相关性较高, 乙酸比例(r=0.77)、丁酸比例(r=-0.85)与第2排序轴相关性较高。因此, 短链脂肪酸对稻秸发酵液的细菌区系多样性存在重要作用。

|
| RF:瘤胃液;RS:稻秸;RS.E:稻秸+乙醇。A:乙酸比例;P:丙酸比例;B:丁酸比例;V:戊酸比例;C:己酸比例 图 5 瘤胃细菌群落与短链脂肪酸产量相关性分析(属水平,典型对应分析) |
体外混合瘤胃细菌培养和每3 d传代一次的培养条件可以保证秸秆的正常发酵, 稻秸体外发酵的SCFA产量因培养时间、发酵底物而不同。艾平等[18]报道, 35℃的中温发酵使稻秸的产酸率最高, 但更高温度(55℃和70℃)的处理不影响SCFA的组成; 牛俊玲等[19]以堆肥样品为材料筛选高效纤维素降解菌, 发现其在发酵前5 d对稻秸的分解活性最高; 张蕴琦提出, 用混合菌系降解水稻秸秆的效果高于单菌株, 且经过驯化后的菌株降解能力增强[20]。短链脂肪酸是瘤胃微生物降解纤维类物质的主要产物, 其中以乙酸、丙酸和丁酸为主, 异丁酸、异戊酸、戊酸和己酸的比例较低。Mirni等[21]和陈安等[22]报道, 稻秸原料接种山羊瘤胃液体外发酵48 h的总挥发性脂肪酸产量为37.3-65.5 mmol/L; 本研究结果表明, 稻秸接种山羊瘤胃液体外发酵72 h的总SCFA产量为78.34 mmol/L, 这一结果与杨红建等[23]研究结果一致, 说明本试验体外发酵稻秸的短链脂肪酸产量在正常范围内。Seedorf等[24]报道, 电子供体是促进SCFA的碳链延伸, 其中乙醇参与乙酸和丁酸发酵, 是己酸合成的重要底物。外源乙醇作为电子供体是影响己酸生产过程中碳链延伸的主要因素[15]。Roghair等[25]在有机废弃物中添加乙醇进行厌氧发酵, 结果表明, 添加乙醇能有效提高己酸的生产速率。本试验中体外稻秸发酵液的己酸产量是瘤胃液的5倍, 添加乙醇的稻秸发酵液己酸产量是瘤胃液的6倍, 该结果表明体外发酵可以显著提高稻秸产己酸的能力。
本试验的Venn图和UPGMA聚类分析都表明, 秸秆发酵液添加乙醇后, 瘤胃细菌多样性发生了变化。拟杆菌门和厚壁菌门被认为是瘤胃中的优势菌门, 成年羊瘤胃中这2个菌门的相对丰度总和>90%[26], 本试验山羊瘤胃液的菌群结构与之相似。瘤胃细菌的菌群相对丰度受日粮结构的影响。范文斌等研究表明, 放牧蒙古羊采食粗纤维高的牧草时, 厚壁菌门相对丰度最高, 拟杆菌门次之[27]。厚壁菌门主要功能是分解纤维类物质, 拟杆菌门的主要功能是分解非纤维类碳水化合物和蛋白质[28]。本试验体外稻秸发酵液的菌门结构发生了显著的改变, 同时添加乙醇后优势菌门相对丰度发生了变化。Zapletalova等[5]双外流连续培养接种瘤胃内容物培养玉米青贮10 d内, 拟杆菌门相对丰度下降, 放线菌门相对丰度增加, 这与本研究结果一致。结合属水平丰度分析和LEfSe分析柱状图结果也可以看出, 秸秆发酵液中添加乙醇显著增加了Biomarker的数量, 其中双歧杆菌(纲、科、属)、放线菌(纲、属)、梭菌芽孢杆菌(纲、目)、脱硫弧菌(纲、目、属)发生了主要的变化。双歧杆菌是肠道中的有益菌, 可抑制病原菌的黏附和侵入, 该菌发酵产生的主要代谢物为有机酸(乙酸为主), 可以通过降低pH值抑制有害菌的增殖[29]; 此外, 有研究表明低乙醇摄入量(占饮水量的10%)可以有助于小鼠肠道双歧杆菌发挥优势菌群的作用, 调节肠道内环境[30]。目前对双歧杆菌在瘤胃内的功能少有报道, 但由于双歧杆菌的主要代谢产物是乙酸, 因此推测该菌的增加可为己酸生产提供更多的底物。克氏梭菌(Clostridium kluyveri)是目前明确报道的可利用乙醇和乳酸产生乙酸, 再进一步利用乙酸延伸碳链产生丁酸和己酸, 因此, 该菌也是研究己酸生产机理的模式菌株[31]。目前对瘤胃内的克氏梭菌的功能研究未见有报道, 但根据本研究结果中梭菌属相对丰度随添加乙醇而增加, 可推测梭菌在瘤胃细菌产己酸中有重要作用。郭威等[32]从窖泥中分离到的放线菌单独培养未检测到己酸产量增加, 但其中8株放线菌与己酸菌共培养发现可有效促进己酸菌生产己酸。吴凌等[33]从奶牛瘤胃中分离出一株产琥珀酸放线杆菌, 并发现该菌与瘤胃液共培养, 显著提高了乙酸、丙酸和丁酸的比例, 降低了乳酸的比例。尽管该研究未检测其它短链脂肪酸, 但可以推测放线杆菌在瘤胃体外发酵中对短链脂肪酸产量有影响。Yang等[34]报道添加乙醇体外培养15 d后显著提高了己酸的产量, 并且脱硫弧菌属为主要的细菌属, Liu等[35]研究也表明脱硫弧菌可利用乙醇作为碳源进行代谢。本试验结果与这些前人结果相似。此外, 从CCA分析中可见, 体外秸秆发酵液的细菌群落多样性受短链脂肪酸的影响。同时, 本试验中添加乙醇使厚壁菌门和放线菌门相对丰度显著升高, 提高了双歧杆菌、放线菌、梭菌和脱硫弧菌属相对丰度, 这可能是导致短链脂肪酸产量发生变化的原因。
4 结论本试验研究了体外瘤胃液与水稻秸秆共培养条件下添加乙醇对短链脂肪酸产量和细菌区系的影响。与体外稻秸发酵液相比, 添加乙醇显著提高了乙酸、戊酸和己酸的比例, 降低了丙酸和丁酸比例; 同时厚壁菌门和放线菌门的相对丰度显著升高。试验结果发现乙醇提高了双歧杆菌、放线菌、梭菌、脱硫弧菌的不同分类水平的相对丰度, 推测这4个菌类在体外发酵水稻秸秆利用乙醇产生己酸上有潜在作用。
| [1] |
农业部科技教育司, 农业部农业生态与资源保护总站.全国农村可再生能源统计汇总表2016[Z].北京: 中国农业出版社, 2016.
|
| [2] |
石祖梁, 贾涛, 王亚静, 等. 我国农作物秸秆综合利用现状及焚烧碳排放估算[J]. 中国农业资源与区划, 2017, 38(9): 32-37. |
| [3] |
毕于运, 高春雨, 王亚静, 等. 中国秸秆资源数量估算[J]. 农业工程学报, 2009, 25(12): 211-217. |
| [4] |
Martínez ME, Ranilla MJ, Tejido ML, et al. Comparison of fermentation of diets of variable composition and microbial populations in the rumen of sheep and Rusitec fermenters. Ⅱ. Protozoa population and diversity of bacterial communities[J]. Journal of Dairy Science, 2010, 93: 3699-3712. DOI:10.3168/jds.2009-2934 |
| [5] |
Zapletalová M, Kašparovská J, Křížová L, et al. Bacterial community dynamics in a rumen fluid bioreactor during in-vitro cultivation[J]. Journal of Biotechnology, 2016, 234: 43-49. DOI:10.1016/j.jbiotec.2016.07.013 |
| [6] |
华金玲, 王立克, 戴四发, 等. 不同处理方法对水稻秸瘤胃降解率的影响[J]. 安徽农学通报, 2013, 19(13): 28-31. DOI:10.3969/j.issn.1007-7731.2013.13.014 |
| [7] |
张轶凤, 杨洪明, 秦廷洋, 等. 常用粗饲料干物质和中性洗涤纤维在瘤胃中降解规律的研究[J]. 饲料工业, 2018, 39(3): 59-64. |
| [8] |
Lin M, Dai XX, Weimer PJ. Shifts in fermentation end products and bacterial community composition in long-term, sequentially transferred in vitro ruminal enrichment cultures fed switchgrass with and without ethanol as a co-substrate[J]. Bioresource Technology, 2019, 285: 121324. DOI:10.1016/j.biortech.2019.121324 |
| [9] |
何迪, 许之扬, 周云龙, 等. 共接种瘤胃微生物和厌氧污泥的水稻秸秆中试厌氧消化系统性能评估[J]. 环境工程学报, 2019, 13(8): 1959-1966. |
| [10] |
Wasewar KL. Reactive extraction :an intensifying approach for carboxylic acid separation[J]. International Journal of Chemical Engineering and Applications, 2012, 3(4): 249-255. |
| [11] |
Zentek J, Buchheit-Renko S, Ferrara F, et al. Nutritional and physiological role of medium-chain triglycerides and medium-chain fatty acids in piglets[J]. Animal Health Research Review, 2011, 12: 83-93. DOI:10.1017/S1466252311000089 |
| [12] |
Serhan M, Mattar J, Debs L. Concentrated yogurt(Labneh) made of a mixture of goats' and cows' milk :physicochemical, microbiological and sensory analysis[J]. Small Ruminant Research, 2016, 138: 46-52. DOI:10.1016/j.smallrumres.2016.04.003 |
| [13] |
Nagao K, Yanagita T. Medium-chain fatty acids :functional lipids for the prevention and treatment of the metabolic syndrome[J]. Pharmacological Research, 2010, 61: 208-212. DOI:10.1016/j.phrs.2009.11.007 |
| [14] |
Chen WS, Strik DP, Buisman CJN, et al. Production of caproic acid from mixed organic waste :an environmental life cycle perspective[J]. Environmental Science and Technology, 2017, 51(12): 7159-7168. DOI:10.1021/acs.est.6b06220 |
| [15] |
Joanna C, Anna D, Roman Z, et al. Caproic acid production from acid whey via open culture fermentation evaluation of the role of electron donors and downstream processing[J]. Bioresource Technology, 2019, 279: 74-83. DOI:10.1016/j.biortech.2019.01.086 |
| [16] |
Menke K, Raab L, Salewski A, et al. The estimation of the digestibility and metabolizable energy content of ruminant feedingstuffs from the gas production when they are incubated with rumen liquor in vitro[J]. Journal of Agricultural Science, 1979, 93(1): 217-222. DOI:10.1017/S0021859600086305 |
| [17] |
张雪娇, 王立志. 饲粮中性洗涤纤维水平对山羊瘤胃细菌结构及组成的影响[J]. 动物营养学报, 2018, 30(4): 1377-1386. DOI:10.3969/j.issn.1006-267x.2018.04.020 |
| [18] |
艾平, 田启欢, 席江, 等. 稻秸预处理厌氧强化产挥发性脂肪酸研究[J]. 农业机械学报, 2018, 49(3): 309-316. |
| [19] |
牛俊玲, 崔宗均, 李国学, 等. 高效纤维素分解菌复合系的筛选构建及其对秸秆的分解特性[J]. 农业环境科学学报, 2005, 24(4): 795-799. |
| [20] |
张蕴琦, 徐凤花, 张书敏, 等. 水稻秸秆降解菌系的筛选及其菌群组成分析[J]. 江苏农业科学, 2017, 45(8): 257-260. |
| [21] |
Mirni L, Ni N, Tri P, et al. Addition of lignocellulolytic enzymes into rice straw improves in vitro rumen fermentation products[J]. Journal of Applied Environmental Biology Science, 2013, 3(9): 166-171. |
| [22] |
陈安, 吴建平, 周传社, 等. C3和C4植物粗料及底物精粗比对山羊瘤胃体外发酵特性的影响[J]. 动物营养学报, 2015, 27(1): 112-123. |
| [23] |
杨红建, 黎大洪, 谢春元, 等. 阿魏酸酯酶处理对羊草、玉米秸、稻秸及麦秸瘤胃体外发酵特性的影响[J]. 动物营养学报, 2010, 22(1): 207-211. |
| [24] |
Seedorf H, Fricke WF, Veith B, et al. The genome of Clostridium kluyveri, a strict anaerobe with unique metabolic features[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105: 2128-2133. DOI:10.1073/pnas.0711093105 |
| [25] |
Roghair M, Liu Y, Strik D, et al. Development of an effective chain elongation process from acidified food waste and ethanol into n-Caproate[J]. Frontiers in Bioengineering and Biotechnology, 2018, 6(50): 1-11. |
| [26] |
张年, 陈明新, 索效军. 不同饲粮条件下湖北黑头羊瘤胃细菌多样性及群落结构分析[J]. 畜牧与兽医, 2019, 51(5): 17-23. |
| [27] |
范文斌, 李长青, 高仙灵, 等. 基于MiSeq技术的枯草期放牧蒙古羊瘤胃细菌多样性分析[J]. 河南农业科学, 2019, 48(4): 146-149. |
| [28] |
Evans NJ, Brown JM, Murray RD, et al. Characterization of novel bovine gastrointestinal tract treponema isolates and comparison with bovine digital dermatitis treponemes[J]. Aoolied and Environmental Microbiolgoy, 2011, 77: 138-147. |
| [29] |
田芬, 陈俊亮, 霍贵成. 嗜酸乳杆菌和双歧杆菌的主代谢产物分析[J]. 中国食品学报, 2013, 13(6): 220-226. |
| [30] |
范观宇, 贺璐, 郑淘, 等. 酒精摄入量对小鼠肠道微生物、酶活性和血常规的影响[J]. 微生物学通报, 2018, 45(7): 1462-1469. |
| [31] |
丁雪松, 赵辉. 克鲁氏梭状芽孢杆菌的研究进展[J]. 食品工业科技, 2012, 33(15): 401-405. |
| [32] |
郭威, 黄宇, 谢逾群, 等. 促己酸菌产己酸的优良放线菌的筛选[J]. 酿酒, 2016, 43(3): 47-51. |
| [33] |
吴凌, 欧海龙, 夏成, 等. 产琥珀酸放线杆菌的分离鉴定及其对瘤胃微生物发酵的影响[J]. 中国微生态学杂志, 2009, 21(8): 737-742. |
| [34] |
Yang PX, Leng L, Tan GY, et al. Upgrading lignocellulosic ethanol for caproate production via chain elongation fermentation[J]. International Biodeterioration and Biodegradation, 2018, 135: 103-109. DOI:10.1016/j.ibiod.2018.09.011 |
| [35] |
Liu FJ, Zhang GP, Liu SR, et al. Bioremoval of arsenic and antimony from wastewater by a mixed culture of sulfate-reducing bacteria using lactate and ethanol as carbon sources[J]. International Biodeterioration and Biodegradation, 2018, 126: 152-159. DOI:10.1016/j.ibiod.2017.10.011 |