




光照强度是影响植物生存、生长发育及分布的重要生态因子[1-3]。光照过强或过弱均对植物造成显著影响, 且不同的植物对光照的需求也有所不同, 因此, 需针对不同植物探究其最适光照区间。磷是植物生长发育不可缺少的大量元素, 其在植物信号传导、能量传递、光合作用和呼吸作用等重要生命过程中起关键作用, 同时磷素也是生物体内核酸、蛋白质、磷脂等生命大分子的重要组成元素[4-7]。金荞麦(Fagopyrum dibotrys(D.Don)Hara), 又名野荞麦、天荞麦、苦荞麦, 是我国Ⅱ级重点保护野生植物。金荞麦根茎可入药[8], 现代中药药理研究发现, 金荞麦可提高机体免疫力和自我防御能力, 在临床上可用于治疗糖尿病、高脂血症、风湿病及癌症等多种疾病[9-10]。随着金荞麦营养价值和药用功能的不断挖掘, 野生金荞麦资源遭到破坏式开采, 濒临灭绝, 产量锐减, 因此, 开展金荞麦人工种植技术研究, 通过人工栽培来提高金荞麦产量及品质意义重大。
植物能通过改变自身的生长状况、形态指标(如根长、株高等)、生物量分配及生理响应等策略来适应光环境的变化[11-14]。刘从等[2]研究发现光照减弱会抑制木本植物幼苗基茎的增长, 但会促进其株高和叶面积的增加; Guo等[15]研究发现遮光34%可以显著促进三角枫生物量积累, 但遮光90%会显著抑制其生物量。对植物而言, 缺磷会使植物电子传递速率(Electro transport rate, ETR)等荧光参数显著降低[6, 16], 相反, 施加磷肥则可以促进植物生长、光合作用, 进而增加产量, 同时也是提高作物品质的重要手段[17-18]。
关于金荞麦与环境因子的研究主要集中在遮阴[19-20]、干旱[21-22]等单因素对其生理生态的影响, 但针对不同光照与磷素协同作用的研究未见报道。
本研究设置不同梯度的光、磷处理, 研究不同光照强度及磷素添加对金荞麦形态、生物量及叶绿素荧光参数的影响, 以及不同光照和施磷处理间的交互作用对金荞麦生长及叶绿素荧光参数的影响, 为种植金荞麦时光照与磷的设置奠定理论基础。
1 材料与方法 1.1 材料试验于2017年在贵州大学生命科学学院实验基地进行, 供试土壤测量后含磷量为0.546 g/kg, 供试金荞麦幼苗购自贵州昌昊金煌中药材有限公司。供试磷源为过磷酸钙, 由天津市科密化学试剂有限公司提供。
1.2 方法 1.2.1 试验设计2017年6月25日将金荞麦幼苗移栽于塑料花盆(内口径×高度为18 cm×23 cm)中。待金荞麦长势稳定后, 选择长势一致的幼苗进行试验处理, 采用黑色聚乙烯遮阳网进行遮阴处理, 遮阴度分别为0(不遮阴, L1)、40%-45%(轻度遮阴, L2)和90%-95%(重度遮阴, L3), 同时每个遮阴处理下设置2个施加过磷酸钙处理(施磷), 施加量分别为0(不施磷, P0)和14.4 g/pot(施磷, P1), 因此, 共L1P0、L1P1、L2P0、L2P1、L3P0、L3P1 6个处理, 每个处理10个重复, 试验过程中保持处理外的其他环境条件一致。试验处理从2017年7月12日持续到2017年9月20日。
1.2.2 测定指标生长指标测定:使用直尺测定株高(Height, H)、总根长(Root length, RL)、叶长(Leaf length, LL)和叶宽(Leaf width, LW), 并统计金荞麦的叶片数(Number of leaves, NOL); 用数显游标卡尺测量叶厚(Leaf thickness, LT)。使用扫描仪测定叶片单叶面积(Single leaf area, SLA), 将金荞麦植株于80℃烘箱烘干至恒重后, 用天平称量金荞麦的叶生物量(Leaf biomass, LB)、茎生物量(Shoot biomass, SB)和根生物量(Root biomass, RB), 并计算总生物量(Total biomass, TB)。
叶绿素及叶绿素荧光指标:使用SPAD-502叶绿素仪测量叶绿素含量(Chlorophyll content, Chl), 使用MINI-PAM2.0测定光化学淬灭系数(Photochemical quenching coefficient, qP)、电子传递效率(Electron transport rate, ETR)、实际光合效率(Yield, YII)、非调节性能量耗散的量子产量(Quantu production of non-regulatory energy dissipation, Y(NO))。
1.2.3 数据统计采用SPSS 22.0对各指标进行方差分析和多重比较, 用Duncan法进行在0.05水平显著性检验, 用Excel 2010进行绘图。
2 结果 2.1 遮阴和施磷对金荞麦形态及生物量的影响由表 1知, 遮阴显著影响金荞麦的根生物量, 对其余指标均表现为极显著影响; 施磷对叶厚和根长的影响不显著, 但显著影响金荞麦的叶片数, 对其余指标均呈现极显著影响; 遮阴和施磷交互作用下, 并未对叶厚和株高产生显著影响, 但显著影响其根生物量和叶生物量, 对其余指标均表现为极显著影响。
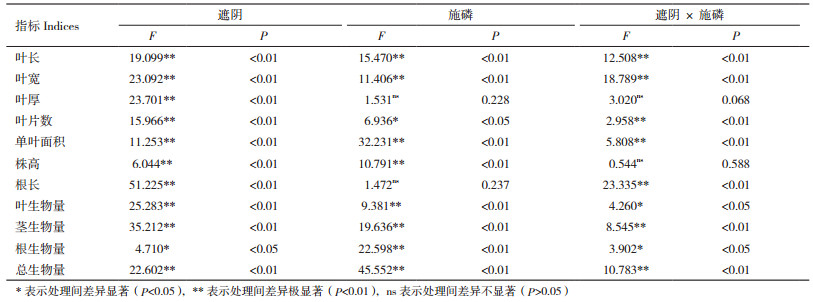
由表 2可知, 在P0处理中, 随遮阴度的不断增加, 金荞麦的叶长、叶宽、叶片数、单叶面积、根长呈先增加后降低的趋势, 但叶长、叶宽和叶面积均大于对照(L1 P0, 下同), 株高呈持续增加, 显著降低叶厚。在L1处理下, 与L1P0相比, L1P1显著增加各形态指标。与对照相比, 遮阴和施磷交互作用(L2P1、L3P1, 下同)显著增加叶长、叶宽、叶面积和株高, 其中, 以L3P1处理最为显著; 叶片数和根长在L2P1处理下较对照也显著增加, 在L3P1处理下不显著降低。这说明单独遮阴有利于金荞麦叶面积增加和株高伸长, 单独施磷对各形态指标均有促进作用, 在轻度遮阴和施磷交互作用下对株高和根长的促进作用最大。
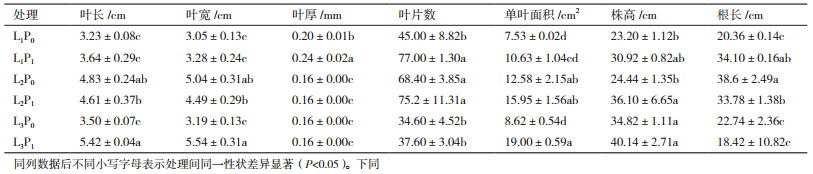
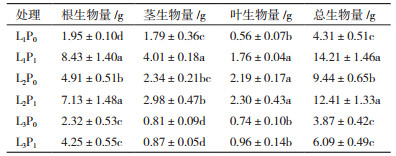
由表 3可知, 在P0处理中, 随遮阴度的增加, 金荞麦生物量积累均呈现出先增加后降低的趋势。在L1处理下, 与L1P0相比, L1P1增加了金荞麦各部分生物量及总生物量积累。与对照相比, 交互作用下金荞麦的生物量积累变化趋势为先增加后降低, 在L2P1处理时均显著增加。说明轻度遮阴有利于增加金荞麦生物量积累, 施磷也促进了金荞麦生物量的积累, 且轻度遮阴和施磷作用时促进作用最大。
由图 1可知, 在P0处理下, 金荞麦的叶绿素含量随遮阴度增大而增加, 且L3P0较L1P0显著增加叶绿素含量。L1处理下, 施磷虽降低金荞麦的叶绿素含量, 但并不显著。与对照相比, 交互作用显著增加叶绿素含量, 且随遮阴度增加而增加。说明施磷虽轻微抑制叶绿素含量增加, 但与遮阴的交互作用缓解了其抑制作用。

|
| 图中不同小写字母表示处理间差异显著(P < 0.05)。下同 图 1 不同光照和磷肥添加下金荞麦叶绿素含量变化 |
由图 2可知, 在P0处理中, 遮阴度增加显著增加金荞麦的YII, 显著降低qP、ETR和Y(NO), 随遮阴度增加影响作用越大。在L1处理下, L1P1较L1P0显著降低金荞麦的ETR和Y(NO), 显著增加qP, 略微增加YII。交互作用对金荞麦qP、YII、ETR和Y(NO)的影响作用与单独遮阴处理的影响作用一致。

|
| 图 2 不同光照和磷肥添加下金荞麦叶绿荧光参数变化特征 |
植物形态特征是植物适应环境的外在表现, 植物会通过改变形态特征来适应环境变化[23], 前人研究发现金荞麦对光照强度变化极敏感, 刘云等[24]研究发现遮阴增加金荞麦的株高、叶长、叶宽、叶面积, 本研究发现, 相比对照和重度遮阴, 轻度遮阴下金荞麦根长、叶片数、叶长、叶宽、叶片数和单叶面积均增加, 说明轻度遮阴较利于金荞麦形态生长。此外, 本研究还发现遮阴后叶厚降低, Sun等[25]认为降低叶厚后能增加植物捕获光能能力并能提高光能利用率, 这说明金荞麦能通过改变自身结构特点来适应不同光环境。
施加磷肥是改善土壤营养不足的重要措施, 缺乏磷肥后植株会出现生长缓慢, 矮小, 地上地下比例失调等现象[4-5, 26-27], 佟静等[28]研究发现适宜的磷素添加显著提高一串红的株高、叶面积等形态指标, 本研究中发现全光下施加磷肥结果与其一致。本研究还发现交互作用下对金荞麦叶长、叶宽、叶面积和株高的促进作用均比单一处理时高, 说明两者交互较利于金荞麦叶片变大及株高伸长, 但轻度及重度遮阴与施磷的交互作用却降低了根长, 这可能是因为本研究是在土壤表面施加磷肥, 根系为获得最大磷元素, 减少了根系长度的投入。本研究发现轻度遮阴及施磷处理下金荞麦叶面积、叶片数、根长及分枝数最大, 从形态特征来看, 轻度遮阴及施磷环境更适合金荞麦形态构成。
3.2 遮阴和施磷处理对金荞麦生物量分配的影响生物量是植物能量合成、同化、转移、贮藏的基本形式, 植物体的构件性状差异最终体现在植株各部分的生物量分配格局, 而各部分生物量通常与光照、营养及生物等环境条件的变化有密切关系[23-24, 29-30]。本研究中发现, 不施磷处理下轻度遮阴处理使金荞麦的根、茎、叶及总生物量最大, 重度遮阴后较轻度遮阴降低, 这与前人研究结果一致[24], 说明适度遮阴对金荞麦的生物量积累具有重要意义。不遮阴处理下, 施加磷肥后各部分生物量及总生物量积累均显著增加。本研究还发现轻度遮阴和施磷交互作用较对照显著增加各部分及总生物量积累, 说明轻度遮阴与施磷交互作用有利于金荞麦生物量积累。并且在重度遮阴和施磷交互下, 茎生物量较对照降低, 这说明磷肥的增产作用需要和适当的光照耦合才能达到较好的效果。以上研究结果表明:金荞麦可依据光照程度采取不同的生长策略, 在不遮阴情况下进行施磷可达最佳效果。
本研究还发现, 较对照而言, 单独的施磷、遮阴和交互作用均能显著提高金荞麦的根生物量, 施磷、轻度遮阴、及轻度遮阴与施磷的交互作用均较对照提高金荞麦的茎生物量。金荞麦的根、茎可入药[8], 由本研究结果可知:不遮阴处理下进行施磷处理可最大限度地提高金荞麦的根、茎生物量, 从而提高其经济产量。
3.3 遮阴和施磷处理对金荞麦叶绿素含量及叶绿素荧光参数的影响本研究发现随遮阴强度增加, 金荞麦叶片叶绿素含量逐渐增加, 说明金荞麦在弱光环境下会加快合成叶绿素以来适应不利环境, 在遮阴90%时表现更显著, 与金荞麦适应性极强的生态习性相符[19], 施加磷肥后, 金荞麦叶绿素含量增加, 这与前人在其他植物上的研究结果一致[16], 这说明遮阴和施磷交互作用下有利于金荞麦叶绿素含量的增加。
qP的大小反映了PSII光合色素所吸收用于光化学电子份额的光能多少[31]。本研究发现, 遮阴显著降低qP和ETR, 说明此时PSII开放程度降低, 电子传递效率降低, 用于光化学电子份额降低, 从而降低光合器官损伤[32], 是金荞麦适应暗光环境的策略。磷素是参与光电子传递、光合产物形成以及卡尔文循环等生理生化反应过程的核心元素, 对植物光合作用有着极其重要的影响[33]。YII是光系统Ⅱ的实际光合量子产量(即实际光合效率), 反映叶绿素吸收的光量子用于光学过程所占的比例。Y(NO)表示因PSII反映中心关闭引起的以热和荧光的形式耗散掉的过剩光能所占的比例, 是光伤害的重要指标, 其值升高表明植物受到损伤[34]。本研究发现单独施磷处理或遮阴处理均降低Y(NO), 增加YII, 这表明施磷和遮阴单独作用时均降低了金荞麦的光伤害程度, 从而促进金荞麦的实际光合效率, 且此时的叶绿素含量显著增加(图 1), 金荞麦通过增加光合作用所需色素来缓解电子传递效率的降低带来的负效应。
4 结论遮阴有利于金荞麦形态生长及生物量积累, 提高其叶绿素含量, 并能够更好地保护光反应系统。施磷促进金荞麦形态生长及生物量积累, 显著增加金荞麦的叶片数。不同程度的遮阴和施磷对金荞麦的影响不一, 其中遮阴40%-45%和施磷的交互作用对金荞麦的促进作用较明显。
| [1] |
Gao B, Xia S, Lei X, et al. Combined effects of different nitrogen sources and levels and light intensities on growth and fatty acid and lipid production of oleaginous eustigmatophycean microalga Eustigmatos cf. polyphem[J]. Journal of Applied Phycology, 2017, 1-15. |
| [2] |
刘从, 田甜, 李珊, 等. 中国木本植物幼苗生长对光照强度的响应[J]. 生态学报, 2018, 38(2): 518-527. |
| [3] |
Rozendaal DMA, Hurtado VH, Poorter L. Plasticity in leaf traits of 38 tropical tree species in response to light; relationships with light demand and adult stature[J]. Functional Ecology, 2010, 20(2): 207-216. |
| [4] |
沙海宁, 孙权, 李建设, 等. 不同施磷量对设施番茄生长、产量的影响及最佳施用量研究[J]. 长江蔬菜, 2010(20): 59-62. |
| [5] |
余爱, 杨帆, 张宇, 等. 不同施磷浓度对柱花草和黑籽雀稗根系分布的影响[J]. 草业学报, 2011, 20(3): 219-224. |
| [6] |
侯云鹏, 杨建, 孔丽丽, 等. 不同施磷水平对春玉米产量、养分吸收及转运的影响[J]. 玉米科学, 2017, 25(3): 123-130. |
| [7] |
高志英, 杨永辉, 张兆沛. 磷肥用量对大豆遮复光后干物质与磷素积累特征的影响[J]. 西北农业学报, 2018, 27(2): 194-202. |
| [8] |
谭露霖, 艾蓉, 罗文菊, 等. 金荞麦的化学成分、生理功能及在畜禽养殖中的应用研究进展[J]. 贵州畜牧兽医, 2016, 40(3): 64-66. DOI:10.3969/j.issn.1007-1474.2016.03.026 |
| [9] |
吕桂兰, 张荫麟. 金荞麦引种栽培与其产量和有效成分含量[J]. 中国兽药杂志, 1995(4): 19-22. |
| [10] |
张京, 况燚, 刘力, 等. 金荞麦块根化学成分的研究[J]. 中草药, 2016, 47(5): 722-725. |
| [11] |
程亚娇, 谌俊旭, 王仲林, 等. 光强和光质对大豆幼苗形态及光合特性的影响[J]. 中国农业科学, 2018, 51(14): 2655-2663. DOI:10.3864/j.issn.0578-1752.2018.14.003 |
| [12] |
Lopez G, Boini A, Manfrini L, et al. Effect of shading and water stress on light interception, physiology and yield of apple trees[J]. Agricultural Water Management, 2018(210): 140-148. |
| [13] |
姜宗庆, 李成忠, 汤庚国. 薄壳山核桃光合生理特性对磷素响应的研究[J]. 北方园艺, 2018(1): 19-22. |
| [14] |
颉洪涛, 虞木奎, 成向荣. 光照强度变化对5种耐阴植物氮磷养分含量、分配以及限制状况的影响[J]. 植物生态学报, 2017, 41(5): 559-569. |
| [15] |
Guo X, Guo W, Luo Y, et al. Morphological and biomass characteristic acclimation of trident maple(Acer buergerianum Miq.)in response to light and water stress[J]. Acta Physiologiae Plantarum, 2013, 35(4): 1149-1159. DOI:10.1007/s11738-012-1154-0 |
| [16] |
韩燕青, 刘鑫, 胡维平, 等. 磷和CO2浓度变化对苦草光合生理的影响[J]. 水生生物学报, 2017, 41(5): 1097-1105. |
| [17] |
袁继存, 赵德英, 徐锴, 等. 不同供磷水平对富士苹果光合及叶绿素荧光特性的影响[J]. 中国南方果树, 2017, 46(6): 112-114. |
| [18] |
陈钢.磷水平对西瓜产量、品质、养分吸收及幼苗耐冷性影响的研究[D].武汉: 华中农业大学, 2008.
|
| [19] |
吴惠群, 李光德. 金荞麦生态环境研究[J]. 云南师范大学学报:自然科学版, 1994, 14(4): 102-109. |
| [20] |
张益锋, 何平, 李桂强, 等. 光强对金荞麦幼苗部分生理指标和生物量的影响[J]. 西南大学学报:自然科学版, 2010, 32(4): 6-11. |
| [21] |
张益锋, 何平, 李桂强, 等. 不同施水处理对金荞麦形态和生物量分配的影响[J]. 中草药, 2009, 40(9): 1456-1459. DOI:10.3321/j.issn:0253-2670.2009.09.038 |
| [22] |
赵丽丽, 王普昶, 陈超, 等. 持续干旱对金荞麦生长、生理生态特性的影响及抗旱性评价[J]. 草地学报, 2016, 24(4): 825-833. |
| [23] |
Sevillano I, Short I, Grant J, et al. Effects of light availability on morphology, growth and biomass allocation of Fagus sylvatica, and Quercus robur, seedlings[J]. Forest Ecology & Management, 2016, 374: 11-19. |
| [24] |
刘云.不同条件下金荞麦(Fagopyrum dibotrys(D. Don)Hara)形态可塑性、生理反应及生物量分配[D].重庆: 西南大学, 2006.
|
| [25] |
Sun XY, Lu ZH, Li PH, et al. Ecological adaptation of Eupatorium adenophorum populations to light intensity[J]. Journal of Forestry Research, 2006, 17(2): 116-120. DOI:10.1007/s11676-006-0027-z |
| [26] |
陈思宁, 黄永芳, 邓建忠, 等. 磷元素对受铝胁迫的高州油茶幼苗光合作用的影响[J]. 经济林研究, 2017, 35(3): 115-120. |
| [27] |
李堆淑, 冀玉良. 细黄链霉菌与氮磷钾肥配施对桔梗幼苗的影响[J]. 广西林业科学, 2018, 47(2): 155-158. DOI:10.3969/j.issn.1006-1126.2018.02.007 |
| [28] |
佟静, 李素艳, 孙向阳, 等. 供磷水平对一串红橙香公主生长及根系发育的影响[J]. 浙江农业学报, 2018, 30(3): 386-392. DOI:10.3969/j.issn.1004-1524.2018.03.05 |
| [29] |
Williams S, Adey W, Halfar J, et al. Effects of light and temperature on Mg uptake, growth, and calcification in the proxy climate archive Clathromorphum compactum[J]. Biogeosciences Discussions, 2018, 15(19): 5745-5759. DOI:10.5194/bg-15-5745-2018 |
| [30] |
Del Valle JC, Buide ML, Whittall JB, et al. Phenotypic plasticity in light-induced flavonoids varies among tissues in Silene littorea, (Caryophyllaceae)[J]. Environmental & Experimental Botany, 2018, 153: 100-107. |
| [31] |
李志博, 魏亦农, 张荣华, 等. 棉花不同叶位叶绿素荧光特性初探[J]. 棉花学报, 2005, 17(3): 189-190. DOI:10.3969/j.issn.1002-7807.2005.03.014 |
| [32] |
司继播, 孙明, 刘良云.叶绿素荧光分析技术综述[C]// 2007年中国农业工程学会学术年会论文摘要集. 2007.
|
| [33] |
王俊秀, 周扬颜, 韩潇, 等. 超表达杨树SBPase基因促进拟南芥光合作用及营养生长[J]. 北京林业大学学报, 2018, 40(3): 26-33. |
| [34] |
Klughammer C, Schreiber U. Complementary PS Ⅱ quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the Saturation Pulse method[J]. Pam Application Notes, 2008, 1: 27-35. |