




烤烟上部叶约占整株烟叶叶数的33%,占烟叶总产量的40%左右,对烤烟总产质量有巨大贡献[1]。优质的上部烟叶具有高烟碱含量和香气量,在现代低焦油烤烟型和混合型卷烟叶组配方中占重要地位,对卷烟的香气和风格也有重要影响[2]。目前,我国上部烟叶叶片窄、组织结构致密、僵硬、填充性差,是导致上部叶质量较差、可用性偏低的重要因素[3-7]。因此,促进烤烟上部叶生长能有效提高烟叶的产量和品质。
温度对烤烟上部叶生长发育、品质和产量产值等均具有一定影响。周越等[8]研究发现,夜温升高能使烤烟上部叶叶长和叶宽显著提高,产量产值显著增加。王佩等[9]研究发现,成熟期增加地温会使上部叶多酚和西柏类化合物含量降低,从而影响上部叶品质。聂荣邦等[10]研究发现,对烟株上部叶进行人工遮阴降温处理有利于提高上部叶成熟度,提高上等烟比例。
目前,有关温度对上部叶生长发育影响的机理研究尚少。本研究以云烟116为材料,研究增温对上部叶胞壁羟脯氨酸含量、胞壁过氧化物酶(Peroxidase,POD)活性和吲哚乙酸氧化酶(Indoleacetate oxidase,IAAO)活性的影响以及这些物质与叶片生长的关系。旨在为促进上部叶生长发育,提高烟叶产量产值提供参考。
1 材料与方法 1.1 材料试验于2017年在贵州省毕节市威宁县黑石头镇烟草科技园试验田中进行。试验田海拔高度2 212 m,东经104°01'06″,北纬26°44'45″,属亚热带季风湿润气候区,年均温16℃,年降水量626 mm,无霜期191 d。试验地土壤为黄棕壤,前茬作物菊花,土壤基础肥力指标:全氮1.63 g/kg,碱解氮46.91 mg/kg,速效磷15.39 mg/kg,速效钾82.00 mg/kg,有机质23.13 g/kg,土壤pH 7.59。
供试烤烟品种为当地主栽品种云烟116。云烟116为8610-711与单育2号杂交育成的烤烟纯系品种。
1.2 方法 1.2.1 试验设计设置2个处理:自然常温对照(T1)和平顶后增温处理(T2)。试验采用小区对比设计,每处理重复3次,每处理100株烤烟,行距110 cm,株距60 cm。试验小区设3垄保护行相隔,所有栽培措施与环境调控措施严格保持一致。
增温处理各小区于7月10日搭设大棚,大棚面积约为63.3 m2,棚高约为2.2 m。大棚用不锈钢管搭建固定,采用透光性极强的透明塑料专用膜覆盖(青州市鲁冠塑料厂生产),减少光质、光强等对试验结果的干扰。为降低试验过程中降雨对试验的影响,对照也设置相应防雨棚(防雨棚材料与大棚相同)。为保证烟株生长用水,每隔5 d给烟株浇水。每个棚内放置一台双金属自动记录温度仪进行温度记录。
1.2.2 测定项目及方法于7月10日烟株平顶始取样,并挂牌做标记定好叶位(自下向上数第17片叶),随后每隔5 d取1次样,共取5次,样品液氮保存备用。
每次取样时用卷尺测量已定位的第17片叶的叶长和叶宽,每次测量10株,用长宽法计算叶面积,叶面积=0.634 5×(叶长×叶宽);利用液质联用方法测定烟叶样品中的羟脯氨酸(Hydroxyproline)含量;参照Maehly和Chance(1954)[11]的方法测定胞壁过氧化物酶活性;利用吲哚乙酸氧化酶试剂盒测定吲哚乙酸氧化酶活性;烘烤后烟叶按照国家烤烟分级标准GB2635-1992进行分级,按烟叶收购价格计算产值、均价与上等烟比例。
1.2.3 统计分析试验数据采用Excel 2003进行整理和制图,采用SPSS 22.0进行统计分析,Duncan新复极差法进行多重比较。
2 结果 2.1 不同处理下温度变化分析由图 1可看出,从7月10日至7月30日,平顶后大棚增温处的棚内平均气温为25.1℃,而防雨棚内的平均气温仅为19.4℃。参照毕节市气象台发布的7月份温度数据,发现7月10日至7月30日防雨棚内的日平均温度与自然常温几乎相同,可以认为大棚内平均气温较自然常温高5.7℃。说明搭建大棚可有效提升烤烟生长的环境温度。

|
| 图 1 不同处理下日平均温度变化示意图 |
由图 2可看出,2个处理上部叶的羟脯氨酸含量随时间推移均呈降低趋势,但较T1处理相比,T2处理的羟脯氨酸含量降低速率更快,从7月15日至7月30日,T2处理上部叶的羟脯氨酸含量显著低于T1处理(P < 0.05)。说明平顶后适当增温可有效降低烟草上部叶叶片细胞壁的羟脯氨酸含量。

|
| 图 2 不同处理下上部叶的羟脯氨酸含量变化 图上不同字母表示不同处理在P < 0.05水平差异显著。下同 |
由图 3可看出,随时间的推移,T1处理的上部叶POD活性呈先略升高后降低然后再升高的变化趋势,而T2处理的上部叶POD活性则呈持续降低趋势,从7月15日至7月30日,T2处理的上部叶POD活性显著低于T1处理(P < 0.05)。说明平顶后适当增温能够降低上部叶叶片细胞壁的POD活性。

|
| 图 3 不同处理下上部叶的POD活性变化 |
由图 4可看出,2个处理上部叶的IAAO活性随时间推移均呈降低趋势,但与T1处理相比,T2处理的上部叶的IAAO活性降低速率更快。从7月15日至7月30日,T2处理上部叶的IAAO活性显著低于T1处理(P < 0.05)。说明平顶后适当增温能够降低上部叶片的IAAO活性。

|
| 图 4 不同处理下上部叶叶片的IAAO活性变化 |
由图 5可看出,2个处理的上部叶叶长均随时间的推移而增长,且T2处理增加的速率大于T1处理。从7月15日至7月30日,T2处理的上部叶叶长始终大于T1处理,且差异显著(P < 0.05)。

|
| 图 5 不同处理下烤烟上部叶叶长变化 |
由图 6可看出,2个处理的上部叶叶宽也随时间的推移而增宽,7月15日之前,T2处理的增长速率较快,7月15日之后,两处理的增长速率相近。从7月15日至7月30日,T2处理的上部叶叶宽始终大于T1处理,且差异显著(P < 0.05)。7月30日时,T2处理的上部叶叶长、叶宽和叶面积分别较T1处理高8.6%、3.9%和12.9%。说明平顶后适当增温能够有效促进烤烟上部叶的生长发育。

|
| 图 6 不同处理下烤烟上部叶叶宽变化 |
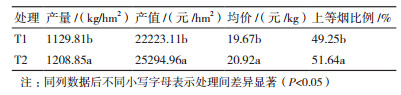
由表 1可知,在平顶后增温条件下,烤烟上部叶的经济性状指标均显著提高。与T1处理相比,T2处理的产量、产值、均价和上等烟比例分别提高7.1%、13.8%、6.4%和4.9%。
本研究以烤烟品种云烟116为材料,测定平顶后不同温度下上部叶胞壁羟脯氨酸含量、胞壁过氧化物酶活性和吲哚乙酸氧化酶活性。结果表明,在平顶后增温条件下,胞壁过氧化物酶活性、吲哚乙酸氧化酶活性和胞壁羟脯氨酸含量均显著降低,上部叶叶长、叶宽和叶面积均显著提高。羟脯氨酸是伸展蛋白重要的组成物质,而伸展蛋白作为植物细胞壁结构蛋白,在初生壁中约占壁干质量的1%-15%,是影响细胞壁伸展性的重要因素之一[12]。而植物细胞伸长和扩大在很大程度上都取决于细胞壁的伸展性。因此,羟脯氨酸影响着植物细胞的生长。Cleland等[13]研究发现,豌豆上胚轴从伸长到停止这段时间内,细胞壁中的羟脯氨酸含量反而增加。Roberts等[14]研究发现,用多肽羟化酶阻遏物处理过的洋葱根伸展蛋白水平降低,导致根细胞伸长。还有研究发现,在培养燕麦胚芽鞘的介质中加入脯氨酸,能抑制羟脯氨酸的形成,导致细胞伸展性增加[15]。这些研究表明伸展蛋白能够抑制细胞的伸展,伸展蛋白的水平随羟脯氨酸含量增加而提高。本研究发现经平顶后增温处理后,细胞壁中的羟脯氨酸含量降低,而上部叶叶长、叶宽和叶面积显著增加,有利于上部叶的生长,与前人研究结果相符。
伸展素(Expansin)是植物细胞生长期间释放的一种能使细胞壁松弛的蛋白质。有研究表明,伸展素中大多羟脯氨酸(Hyp)残基通过Hyp-O-Ara纽带与壁多糖相联,形成网络,限制细胞生长(纽带说)[16]。但也有研究发现,伸展素多肽链间相邻的异二酪氨酸(Idt)残基通过二苯基醚键两两相连形成网络,限制细胞生长(自身交联说)[17]。而在这两种途径中,胞壁POD对Hyp和Idt的形成都起着重要作用。研究发现,胞壁POD能够催化伸展素中的脯氨酸(Pro)羟化,形成羟脯氨酸,使羟脯氨酸与脯氨酸之比增大,从而不利于细胞生长。POD也能使相邻酪氨酸(Tyr)氧化形成Idt,限制细胞生长[18]。黄海等[19]研究了伸展蛋白与烟草叶细胞扩大生长的关系,结果表明,生长缓慢的圆片细胞壁强脯氨酸与脯氨酸的比值高于迅速生长的圆片。本研究发现,在增温处理下,胞壁POD活性降低,羟脯氨酸含量减少,可能是因为POD活性降低,催化Pro羟基化能力变弱,羟脯氨酸与脯氨酸比值变小,导致叶片伸展。当然,也不排除自身交联说。在增温条件下,胞壁POD活性降低,可能使其氧化Tyr形成Idt的能力变弱,通过自身交联形成的网络就变少,从而促使细胞伸展。但也有研究表明,伸展素能参与植物细胞壁酸性扩张。Cosgrove实验室分别从黄瓜下胚轴细胞壁和玉米花粉中分离出α-伸展素和β-伸展素,二者均能恢复细胞壁酸性扩张[20],并据此推测伸展素在纤维素微纤丝表面扩散,以一种可逆的方式作用于与纤维素微纤丝表面紧密结合的基质聚合物,使多聚体网络之间的氢键断裂,膨压驱使纤维素微纤丝与半纤维素相互之间滑动,从而引起细胞壁伸长[21]。本研究结果与其相反,可能是试验材料和试验环境不同导致的。由于本研究并未对Hyp、Tyr、Idt等物质进行测定,关于伸展素在POD的作用下抑制叶片生长的具体途径还需进一步研究。
Lamport等[22]在研究POD的生理功能时发现,过氧化物酶还能通过氧化生长素的方式影响细胞生长。IAA作为生长素中最重要的一种内源激素,与细胞分裂和扩大更是密切相关[23]。而IAAO和POD作为IAA分解过程中的关键酶,均能氧化分解IAA,调节植物生长[24]。本研究发现,在平顶后增温处理下,IAAO和POD活性降低,上部叶叶长、叶宽和叶面积增大。这是因为当IAAO和POD活性降低时,氧化分解IAA的能力变弱,因此,与对照相比,被分解的IAA较少,从而促进细胞生长,有利于上部叶开片。但关于IAA促进上部叶细胞生长的最适浓度还有待进一步研究。
4 结论平顶后适当增温能够显著降低烤烟上部叶叶片POD活性、IAAO活性和羟脯氨酸含量,显著提高上部叶叶长、叶宽和叶面积,有效地促进烤烟上部叶的生长发育。经大棚增温处理后,温度提高了5.7℃,烤烟上部叶叶长、叶宽、叶面积、产量和产值显著增加,较对照相比分别提高了8.6%、3.9%、12.9%、7.0%和13.8%。
| [1] |
许自成, 黄平俊, 苏富强, 等. 不同采收方式对烤烟上部叶内在品质的影响[J]. 西北农林科技大学学报:自然科学版, 2005, 33(11): 13-17. |
| [2] |
宫长荣, 刘霞, 宋朝鹏, 等. 影响烤烟上部叶质量的因素及提高其可用性的措施[J]. 中国农学通报, 2007(3): 103-108. DOI:10.3969/j.issn.1000-6850.2007.03.024 |
| [3] |
梁斌, 蔚应俊, 周应兵. 烤烟上部叶滞销的原因及农业生产对策[J]. 安徽农业科学, 2002, 30(2): 285-286. DOI:10.3969/j.issn.0517-6611.2002.02.061 |
| [4] |
纪成灿, 王胜雷, 许锡祥. 提高上部叶可用性和降低上部叶比例的农业措施[J]. 中国烟草科学, 2001, 22(4): 19-22. DOI:10.3969/j.issn.1007-5119.2001.04.013 |
| [5] |
唐经祥, 何厚民, 江理论.关键农业技术措施对烤烟上部叶外观质量及经济性状的影响[A]//陈江华.中国烟叶学术论文集[C].北京: 科学技术出版社, 2004: 107-109.
|
| [6] |
刘碧荣.五项措施对烤烟上、中部叶烟碱及其它主要化学成分的影响[D].贵阳: 贵州大学, 2006. http://d.wanfangdata.com.cn/Thesis/Y960865
|
| [7] |
邱标仁, 林桂华. 提高龙岩烟区上部叶可用性的途径[J]. 中国烟草科学, 2000, 21(2): 16-18. DOI:10.3969/j.issn.1007-5119.2000.02.004 |
| [8] |
周越, 范幸龙, 周冀衡, 等. 夜温升高对云南省高海拔烤烟理化特性及产、质量的影响[J]. 中国生态农业学报, 2015, 23(1): 61-68. |
| [9] |
王佩, 尤磊, 刘朝科, 等. 成熟期地温对烟叶主要致香物质含量的影响[J]. 安徽农业科学, 2014, 42(20): 6569-6572. DOI:10.3969/j.issn.0517-6611.2014.20.016 |
| [10] |
聂荣邦, 韦建玉, 汪少波, 等. 烤烟上部叶遮荫处理对田间小气候及烟叶质量的影响[J]. 作物研究, 2013, 27(6): 611-613. DOI:10.3969/j.issn.1001-5280.2013.06.20 |
| [11] |
Maehly AC, Chance B. Catalases and peroxidases, part Ⅱ[J]. Methods of Biochemical Analysis, 1954, 1(3): 357-424. |
| [12] |
刘晶晶, 邓泽元. 植物细胞壁中的伸展蛋白[J]. 氨基酸和生物资源, 2003(3): 20-23. DOI:10.3969/j.issn.1006-8376.2003.03.007 |
| [13] |
Cleland R. Hydroxyproline as an inhibitor of auxin-induced cell elongation[J]. Nature, 1963, 200(490): 908. |
| [14] |
Roberts K, Shirsat AH. Increased extensin levels in Arabidopsis affect inflorescence stem thickening and height[J]. Journal of Experimental Botany, 2006, 57(3): 537. DOI:10.1093/jxb/erj036 |
| [15] |
Cleland R, Karlsnes AM. A possible role of hydroxyproline-containing proteins in the cessation of cell elongation[J]. Plant Physiology, 1967, 42(5): 669-671. DOI:10.1104/pp.42.5.669 |
| [16] |
郭联华, 尹佟明, 李淑娴, 等. 植物细胞壁伸展蛋白研究述评[J]. 林业科学, 2010, 46(12): 144-152. DOI:10.11707/j.1001-7488.20101224 |
| [17] |
李雄彪, 杨中汉. 伸展蛋白的结构、功能、交联和生物合成[J]. 植物生理学通讯, 1990(3): 7-13. |
| [18] |
甘苏生, 沈曾佑, 张志良, 等. 黄瓜子叶细胞壁扩大生长机理的研究[J]. 植物学报, 1987(4): 379-387. |
| [19] |
黄海, 沈曾佑, 张志良, 等. 细胞壁中富含羟脯氨酸蛋白质与烟草叶细胞扩大生长的关系[J]. 植物生理学报, 1983(1): 85-92. |
| [20] |
Cosgrove DJ. Loosening of plant cell walls by expansins[J]. Nature, 2000, 407(6802): 321-326. DOI:10.1038/35030000 |
| [21] |
赵庆新, 袁生. 植物细胞壁研究进展[J]. 生物学通报, 2007(7): 8-9. DOI:10.3969/j.issn.0006-3193.2007.07.003 |
| [22] |
Lamport DTA. 13-Structure and function of plant glycoproteins[J]. Carbohydrates Structure & Function, 1980, 67(6): 501-541. |
| [23] |
王家利, 刘冬成, 郭小丽, 等. 生长素合成途径的研究进展[J]. 植物学报, 2012, 47(3): 292-301. DOI:10.3969/j.issn.1005-3395.2012.03.013 |
| [24] |
原牡丹, 侯智霞, 翟明普, 等. IAA分解代谢相关酶(IAAO、POD)的研究进展[J]. 中国农学通报, 2008(8): 88-92. |