




2. 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,银川 750021
2. Key Laboratory for Restoration and Reconstruction of Degraded Ecosystem in Northwest China of Ministry of Education, Ningxia University, Yinchuan 750021
毛乌素沙地地处农牧业交错带,是我国北方重要的生态屏障,植被类型属于草原向荒漠草原过渡的类型[1]。植被类型单一,表层植被以旱生植物为主[2]。其中,油蒿(Artemisia ordosica)是毛乌素沙地最重要的建群植物之一,占沙区总面积31.2%,是毛乌素沙地面积最大的固沙植物群落[3],对维护当地生态系统稳定起着重要作用。由于油蒿根系非常发达,其持续生长对土壤水分消耗十分强烈,土壤干燥化现象普遍发生,出现植被逐渐退化的现象[4]。因此,在植被恢复过程中,必须考虑植物生长与土壤耗水量的关系。
植物的生理指标变化是对植物生长状况的综合反映,植物生理活动受多种环境因素的影响[5]。其中立地因子是一个重要的影响因素,而坡位是一个重要的立地因子,太阳辐射强度和降雨在土壤中的再分配进一步改变土壤含水量,并且会影响植被的分布以及其他生态因子的变化,对植物生长有着重要的影响。20世纪90年代以来,在灌木、乔木中广泛开展了水分胁迫抗性生理评价的研究[6-9]。朱林等[10]利用氧同位素对宁夏盐池不同坡位旱地紫花苜蓿(Medicago sativa)水分来源进行研究,发现紫花苜蓿适宜在西北半干旱地区的低洼滩上种植。王洋等[11]研究不同坡位红砂的生理指标,结果表明,红砂主要通过调节SOD、POD等生理指标来适应坡位的变化。朱志东[12]研究甜橘柚不同坡向、坡位生长和生理指标,发现坡位直接或间接地影响植物生长、营养状况、以及林分发展。刘旻霞等[13]研究了亚高寒草甸植物生理指标对坡向的响应,发现微地形上植物叶片生理特性受环境因子影响显著。
目前有关植物的土壤水分和生理指标研究较多,但对于毛乌素沙地沙生灌丛,特别是其生理指标结合土壤水分研究微地形与油蒿灌木之间的关系还鲜有报道。
本研究以宁夏盐池毛乌素沙地典型沙生灌木油蒿为研究对象,研究处于不同坡位的油蒿,持续干旱胁迫对植物生理指标及土壤水分的影响,探讨随干旱胁迫的延长植物生长与土壤水分消耗之间的相互作用关系,为揭示沙生植物抗旱生理生化机制提供理论依据,为毛乌素沙生植物的保护和修复提供一定的理论依据,为未来环境变化下荒漠植物群落的响应提供理论参考。
1 材料与方法 1.1 材料试验地位于宁夏回族自治区吴忠市盐池县高沙窝乡,地理坐标界于37°02’08”-38°26’58”N和101°03’24”-102°15’35”E,该样地处于毛乌素沙地东南缘,地貌类型以起伏的固定和半固定沙丘为主,区内海拔1 450-1 480 m。该地区属于典型中温带大陆性气候,冬季长,温差大,干燥寒冷,年降水量少并且分布不均,蒸发量较大,年均温度为8.1℃,年均无霜期为165 d。试验地内土壤种类主要是风沙土,pH为8.5-8.8[14],有机质含量低,腐殖质缺乏,土壤肥力极差。植被类型属于草原向荒漠草原过渡的类型,植被种类贫乏,地表植被主要由旱生类型的植物组成。主导的灌木为油蒿(Artemisia ordosica)、杨柴(Hedysarum mongolicum Turez)、中间锦鸡儿(Caragana intermedia Kuang et H.C.Fu),在样地内多呈簇状均匀散布;草本植物以苦豆子(Sophora alopecuroides)、甘草(Glycyrrhiza uralensis Fisch)、中亚白草(Pennisetum centrasiaticum)、赖草(Leymus secalinus)为主。其中,柠条在人工林中有很高的重要值;油蒿在各种天然灌木林中的分布较广,重要值也较高。在整个试验样地的植被类型中,油蒿是主要的建群种和优势种,盖度为50%-80%。试验区降雨量及气温特征如图 1所示。

|
| 图 1 日降雨量及日平均气温特征 |
于2018年7-9月开展野外试验,在固定样地选择2个植被分布几乎一致的沙丘,在其顶部、中部及丘间低地选择油蒿(Artemisia ordosica)灌木植物作为研究对象,每个坡位固定8-10株大小长势及冠幅一致的灌丛,布设大小均为6 m×6 m的3个重复样方。采集叶片时,应选择自然生长状态下新生枝条上的叶片,将刚采集的叶片样品装入液氮罐中,以备测定。用直径5 cm的土钻在每个样方内采用五点法钻取土壤(0-40 cm土层)混匀后分析其化学性质,并用环刀法测定土壤的物理性质。同时,用美国HOBO U30土壤湿度自动记录仪测定土壤水分。
1.2.2 指标测定土壤含水量采用TDR土壤水分探测仪测定[15];采用电导法测定细胞膜透性[16];采用TBA(硫代巴比妥酸)测定MDA含量[17];SOD活性采用核黄素-NBT法测定[18];POD活性采用愈创木酚比色法测定[18]。
1.2.3 数据分析采用Excel 2010软件进行数据整理,用SPSS 25.0统计软件进行显著性分析和相关性分析,利用SigmaPlot 12.5作图并用字母法标记。
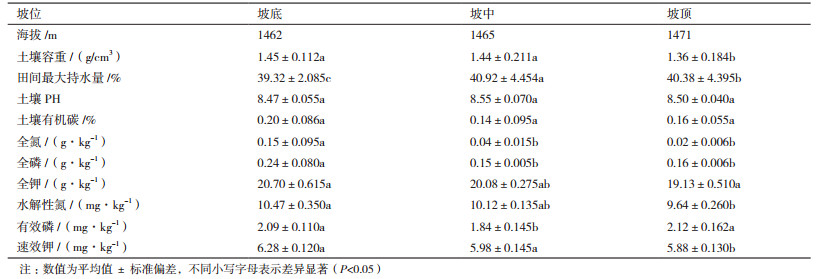
2 结果 2.1 试验区不同坡位土壤理化性质试验区不同坡位土壤理化性质如表 1所示,土壤容重为1.36-1.45 g/cm3;坡底和坡中无显著差异,但显著大于坡顶;田间最大持水量为39.32%-40.92%,不同坡位之间存在显著差异,排序为坡中>坡顶>坡底;不同坡位土壤pH为8.4-8.6,无显著性差异(P < 0.05);土壤养分方面:随着坡位的升高呈逐渐下降趋势。土壤有机碳的变化为坡底>坡顶>坡中,但之间无显著差异;全氮、全钾含量坡底明显大于坡中和坡顶(P < 0.05);水解性氮、速效钾含量坡顶与坡底和坡中有显著差异(P < 0.05);而有效磷含量变化为坡顶>坡底>坡中,坡中和坡底、坡顶之间差异显著(P < 0.05)。
由图 2可以看出,不同时期土壤含水量的变化趋势为7月29日>8月31日>10月4日>9月16日。土壤含水量总体表现为坡底>坡中>坡顶,不同坡位之间差异显著。

|
| 图 2 不同坡位土壤体积含水量的变化 |
由图 3可知,4个采样时期0-300 cm土壤含水量的排序为:7月29日>8月31日>9月16日>10月4日。由于降雨主要集中在7月,故7月土壤含水量最大。随着气温升高,降雨量减少,油蒿对水分的需求也开始增加,土壤含水量开始下降,这也符合土壤水分含量在浅层变化块的特点[19]。7月、8月、9月及10月0-300 cm土壤剖面平均含水量排序均为坡底>坡中>坡顶,分别为23.69%、20.13%和18.84%;22.48%、19.04%和17.52%;19.98%、16.48%和15.24%;20.32%、16.79%和15.58%。坡底和坡中油蒿土壤剖面土壤含水量呈浅层低深层高的趋势。坡中呈浅层及深层低,中间层次较高的趋势。

|
| A:7月29日;B:8月31日;C:9月16日;D:10月4日 图 3 不同时期3个坡位油蒿0-300 cm土层土壤含水量剖面分布 |
由图 4可知,4个采样期不同坡位油蒿叶片细胞膜透性均存在显著差异(P < 0.05)。在7月坡中和坡顶油蒿叶片细胞膜透性无显著差异,但显著低于坡底(P < 0.05);在8月坡底油蒿叶片细胞膜透性显著高于其他2个坡位(P < 0.05),坡顶油蒿叶片细胞膜透性也显著高于坡中(P < 0.05);9月坡中油蒿叶片细胞膜透性显著低于其他2个坡位(P < 0.05),其他两个坡位叶片细胞膜细胞膜透性无显著差异;在10月,坡底和坡顶油蒿叶片细胞膜透性差异不显著,但显著低于坡中(P < 0.05)。

|
| 不同小写字母表示同一时期不同坡位油蒿叶片生理指标差异显著(P < 0.05);不同大写字母表示同一坡位不同时期油蒿叶片生理指标差异显著(P < 0.05)。下同。 图 4 不同时期3个坡位细胞膜透性(x±SD) |
坡中油蒿叶片细胞膜透性在7月、8月及9月都没有显著差异,但显著低于10月(P < 0.05);9月坡底油蒿叶片细胞膜透性显著高于7月和8月(P < 0.05),但与10月无显著差异;坡顶油蒿叶片细胞膜透性在7月、8月及9月有显著差异,排序为9月> 8月> 7月。
2.5 不同坡位对油蒿叶片MDA含量的影响由图 5可知,4个采样期不同坡位油蒿叶片MDA含量均存在显著差异(P < 0.05)。在7月,坡底油蒿叶片MDA含量显著低于坡中和坡顶(P < 0.05),但坡中和坡顶无显著差异;在8月,坡中油蒿叶片MDA含量高于其他2个坡位(P < 0.05),坡底和坡顶油蒿叶片MDA含量无显著差异;9月和10月,3个坡位叶片MDA含量有显著性差异,排序为坡底>坡中>坡顶(P < 0.05)。

|
| 图 5 不同时期3个坡位MDA含量(x±SD) |
坡底4个时期叶片MDA含量变化为9月>10月>8月>7月(P < 0.05);其他2个坡位4个时期的叶片MDA含量都没有显著差异。
2.6 不同坡位对油蒿叶片抗氧化酶活性的影响由图 6可知,将同一坡位不同月之间的叶片POD相比较可以发现:9月坡底油蒿叶片POD高于7月、8月和10月(P < 0.05)。坡中叶片POD不同月无显著差异。坡顶9月叶片POD与10月无显著差异,但显著高于7月、8月(P < 0.05),并且7月和8月也无显著差异。

|
| 图 6 不同时期3个坡位POD活性(x±SD) |
4个时期坡底油蒿叶片POD均显著高于其他2个坡位(P < 0.05)。7月坡中和坡顶油蒿叶片POD无显著差异;8月坡中油蒿叶片POD显著高于坡顶,但9月和10月坡中油蒿叶片POD显著低于坡顶。
由图 7可知,7月、8月及10月不同坡位间油蒿叶片SOD均有显著差异,9月不同坡位之间无显著差异。9月和10月不同坡位间SOD顺序是坡中>坡顶>坡底,但7月和8月不同坡位间SOD是坡中>坡底>坡顶。

|
| 图 7 不同时期3个坡位SOD活性(x±SD) |
不同月同一坡位之间均有显著差异。坡底油蒿叶片SOD的变化趋势为9月>10月>8月>7月(P < 0.05);坡中和坡顶叶片SOD在9月及10月无显著差异,但显著高于7月和8月,其中,7月和8月也无显著差异,SOD的变化趋势为10月>9月>8月>7月(P < 0.05)。
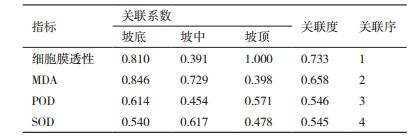
2.7 指标的排序分析由表 2所示,选择油蒿细胞膜透性、MDA、POD和SOD 4个指标,计算关联系数和关联度,依据关联度进行关联排序,影响力大小排序为:细胞膜透性>MDA>POD>SOD。这说明细胞膜透性与坡位的抗旱性关联较大,是衡量不同坡位抗旱性的首要指标。
植物在生长发育过程中随干旱胁迫的变化其生理指标也发生不同程度的变化[20]。本研究结果表明,油蒿在不同坡位生理指标对干旱的响应机制各不相同。已有研究发现植物的细胞膜透性和MDA含量与植物的耐旱性呈负相关[21]。本研究中,3个坡位的细胞膜透性和MDA含量在轻微干旱胁迫下总体呈上升趋势,MDA含量变化和细胞膜透性的变化趋势基本吻合,与植株的抗旱能力呈一定的负相关,细胞膜透性的增幅为坡中 > 坡顶 > 坡底,而MDA含量坡底的上升速度显著大于坡中和坡顶。
研究表明,植物SOD活性在持续干旱胁迫下,会出现先升高后降的趋势。本研究3个坡位SOD活性呈现先升高后降底的趋势,表现为9月>10月>8月>7月,这与之前的研究结果一致[22]。邹原东等[23]发现植物(如金心吊兰)细胞在轻微干旱胁迫下POD和SOD酶活性会增高,但干旱胁迫过高时,POD和SOD酶活性下降。本研究表明,随着干旱胁迫的延长,3个坡位油蒿的POD和SOD酶活性逐渐上升,但当干旱胁迫过大时POD和SOD酶活性下降,这与冯士令等[24]、杨舒贻等[25]的研究结果均相似。不同坡位POD酶活性顺序为:坡底 > 坡中 > 坡顶,由图 2可知,土壤含水量变化为:坡中 > 坡底 > 坡顶,POD随土壤含水量呈现不规则变化,说明油蒿POD酶活性的变化与土壤含水量没有直接关系,可能由于海拔高度引起的其他气象调节变化导致[26]。SOD酶活性在不同坡位7月和8月顺序为:坡中 > 坡底 > 坡顶,与土壤含水量变化一致。9月和10月顺序为:坡中 > 坡顶 > 坡底。
坡位对油蒿的生理指标影响显著,各坡位油蒿生理指标的变化幅度和范围都不同,在干旱胁迫下,各项指标随着胁迫强度的增加而变化,油蒿叶片细胞膜透性、MDA含量、POD和SOD酶活性都随干旱胁迫的增加而增加或者先增加后降低,这些生理指标具有一定的相关动态变化。当植物细胞缺水遭到伤害时,其产生的丙二醛具有较强的细胞毒性,影响植物的正常生长[27]。因此,研究植物的抗旱性对保护生态环境、绿化干旱地区有重要意义。本实验是在野外自然条件下进行的,与室内试验相比,缺乏一定的可控性,而且精确度也有一定的差异,后续研究将增加室内控制实验进行抗旱鉴定。
4 结论坡中土壤含水量最高,各生理指标表现的抗逆性较强,所以油蒿适宜生长在地势居中的缓坡上,以满足植物生长发育的需要并取得较好的生态效益。
| [1] |
陈昌笃. 走向宏观生态学-陈昌笃论文集[M]. 北京: 科学出版社, 2009: 85-115.
|
| [2] |
赵晨光.毛乌素沙地生态承载力及土壤侵蚀特征研究--以乌审旗为例[D].呼和浩特: 内蒙古农业大学, 2014. http://d.wanfangdata.com.cn/Thesis/Y2600797
|
| [3] |
宫攀. 中国土地覆盖分类研究[M]. 北京: 经济科学出版社, 2012.
|
| [4] |
Wang YQ, ShaMA, Liu ZP. Large-scale spatial variability of dried soil layers and related factors across the entire Loess Plateau of China[J]. Geoderma, 2010, 159(1): 0-108. |
| [5] |
刘旻霞. 甘南高寒草甸植物元素含量与土壤因子对坡向梯度的响应[J]. 生态学报, 2017, 37(24): 8275-8284. |
| [6] |
Flexas J, Medrano H. Drought-inhibition of photosynthesis in C3 plants: stomatal and non-stomatal limitations revisited[J]. Annals of Botany, 2002, 89(2): 183-189. DOI:10.1093/aob/mcf027 |
| [7] |
Lawson T, Oxborough K, Morison JI, et al. The responses of guard and mesophyll cell photosynthesis to CO2, O2, light, and water stress in a range of species are similar[J]. Journal of Experimental Botany, 2003, 54(388): 1743-1752. DOI:10.1093/jxb/erg186 |
| [8] |
韩刚, 赵忠. 不同土壤水分下4种沙生灌木的光合光响应特性[J]. 生态学报, 2010, 30(15): 4019-4026. |
| [9] |
张仁和, 郑友军, 马国胜, 等. 干旱胁迫对玉米苗期叶片光合作用和保护酶的影响[J]. 生态学报, 2011, 31(5): 1303-1311. |
| [10] |
朱林, 祁亚淑, 许兴. 宁夏盐池不同坡位旱地紫苜蓿水分来源[J]. 植物生态学报, 2014, 38(11): 1226-1240. |
| [11] |
王洋, 单立山, 白蕾, 等. 不同坡位对红砂生理指标的影响[J]. 水土保持通报, 2017, 37(6): 113-118. |
| [12] |
朱志东. 不同坡向、坡位对甜橘柚生长和生理指标的影响[J]. 食品安全导刊, 2016(3): 126-128. |
| [13] |
刘旻霞, 赵瑞东, 张灿, 等. 亚高寒草甸植物叶片生理指标对坡向的响应[J]. 应用生态学报, 2017, 28(9): 2863-2869. |
| [14] |
苏培玺, 解婷婷, 周柴鹃. 我国荒漠地区C4植物种类分布及其与气候的关系[J]. 中国沙漠, 2011, 31(2): 267-276. |
| [15] |
康洁, 张维江, 李娟. TRIME-T3管式TDR土壤水分测定系统在宁夏泾源地区的标定研究[J]. 宁夏工程技术, 2015, 14(2): 146-148. DOI:10.3969/j.issn.1671-7244.2015.02.012 |
| [16] |
向地英, 张钢, 杨利平. 热激诱导对高温胁迫下百合相对电导率及电阻抗参数的影响[J]. 西北农业学报, 2014, 23(9): 189-195. |
| [17] |
王玲丽, 贾文杰, 马璐琳, 等. 低温胁迫对不同百合主要生理指标的影响[J]. 植物生理学报, 2014(9): 1413-1422. |
| [18] |
朱志国, 季晓莲. 百合鳞茎形成的生理生化研究[J]. 中国农学通报, 2015, 31(10): 138-141. DOI:10.11924/j.issn.1000-6850.casb14100019 |
| [19] |
Yang L, Wei W, Chen L, et al. Response of deep soil moisture to land use and afforestation in the semi-arid Loess Plateau, China[J]. Journal of Hydrology, 2012, 475(6): 111-122. |
| [20] |
罗梓琼, 王慧, 陈铭, 等. 干旱胁迫对4个油茶品种苗木生理生化指标的影响[J]. 经济林研究, 2019(2): 104-113. |
| [21] |
谢志玉, 张文辉, 刘新成. 干旱胁迫对文冠果幼苗生长和生理生化特征的影响[J]. 西北植物学报, 2010, 30(5): 948-954. |
| [22] |
白鹏, 冉春艳, 谢小玉. 干旱胁迫对油菜蕾薹期生理特性及农艺性状的影响[J]. 中国农业科学, 2014, 47(18): 3566-3576. DOI:10.3864/j.issn.0578-1752.2014.18.005 |
| [23] |
邹原东, 韩振芹, 陈秀新, 等. 干旱胁迫对蓝羊草渗透调节物质和抗氧化酶活性的影响[J]. 北方园艺, 2013(23): 71-75. |
| [24] |
冯士令, 程浩然, 李倩倩, 等. 3个油茶品种幼苗对干旱胁迫的生理响应[J]. 西北植物学报, 2013, 33(8): 1651-1657. |
| [25] |
杨舒贻, 陈晓阳, 惠文凯, 等. 逆境胁迫下植物抗氧化酶系统响应研究进展[J]. 福建农林大学学报:自然科学版, 2016, 45(5): 481-489. |
| [26] |
姚毓菲, 邵明安, 贾玉华, 等. 生长年限对苜蓿和柠条光合特征及土壤水分的影响[J]. 干旱地区农业研究, 2019, 37(2): 18-24. |
| [27] |
姜宗庆, 李成忠, 余乐, 等. 干旱胁迫对薄壳山核桃叶片丙二醛含量和3种抗氧化酶活性的影响[J]. 上海农业学报, 2019, 35(1): 7-10. |