




蚜虫的种类很多,有菜蚜、棉蚜、豆蚜、麦蚜等,均属同翅目蚜科[1]。当前辣椒生产中为害的蚜虫主要为桃蚜(Myzus persicae(sulzer))[2]。桃蚜不仅能以成、若蚜群集于寄主的叶背、顶芽、嫩茎等处刺吸汁液,直接造成寄主叶片皱缩或卷曲,被害植株生长缓慢甚至停止等,而且是一种非常重要的媒介昆虫,能传播多种病毒,导致病毒病的发生与流行,造成作物严重减产[3-4]。随着瓜菜生产的快速发展,桃蚜在辣椒、瓜类等作物上的发生与危害也日趋严重,严重制约了瓜菜高效、优质、高产栽培与生产。目前,瓜菜生产中对于桃蚜的防治仍依赖于化学防治,但化学防治过程中存在诸多问题,如施药技术落后带来的农药的有效利用率不足、农药的使用频率及使用剂量不断加大、剧毒、高毒农药的盲目使用等,必将导致害虫产生抗药性、农产品质量安全受到威胁、产地生态环境恶化等严重问题[5]。因此,寻求安全高效的防治措施成为辣椒产业可持续发展亟需解决的重大问题。
利用作物抗虫性防治害虫被认为是最经济、有效、简便的措施[6],其中抗虫品种的筛选和培育是作物抗虫性实际应用的最主要手段[7]。在辣椒抗蚜品种的研究方面,Frantz等[8]从50份辣椒种质中筛选出6份抗桃蚜种质。Bosland和Ellington[9]测定比较了抗蚜和感蚜辣椒种质的生理生化差异,发现非选择性的排拒作用(Nonpreference antixenosis)是辣椒种质产生抗性的主要机制之一。作物品种的抗虫性首先表现在遗传特性上,是作物抗虫遗传特性、害虫危害遗传特性与环境等多种因素相互作用的结果[10]。了解作物的抗虫遗传特性对选育抗虫品种具有重要的指导意义,然而迄今为止尚未见辣椒抗蚜性遗传分析的相关报道。本研究将抗蚜辣椒品种‘猪大肠’与感蚜辣椒品种‘大羊角椒’配置组合,通过F1代抗性鉴定和F2代分离群体抗感分离比例分析,以期明确其抗蚜基因的组成及遗传特点,并测定分离后代辣椒群体在蚜虫为害前后过氧化物酶(Peroxide enzyme,POD)和多酚氧化酶(Polyphenol oxidase,PPO)的活性差异,为抗蚜基因标记奠定基础,为辣椒抗蚜新品种选育提供理论依据。
1 材料与方法 1.1 材料桃蚜采自中国热带农业科学院环境与植物保护研究所害虫研究室盆栽烟草植株。以遗传稳定的抗蚜辣椒品种‘猪大肠’和感蚜辣椒品种‘大羊角椒’为亲本材料。
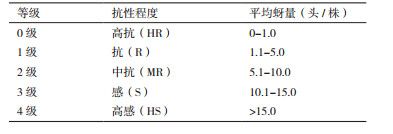
1.2 方法 1.2.1 杂交后代群体的抗蚜性鉴定采用实验室前期建立的辣椒品种温室苗期鉴定方法[11]对正反杂交杂种后代F1、F2进行抗性评价(图 1),将待测品种随机排列,播种于育苗用营养钵中,每品种1行,每行10株,行长60 cm,行距、株距均为6 cm。在辣椒苗4-6叶期进行人工接蚜。方法是剪取布满蚜虫的辣椒叶片,均匀设三点置于待测苗的行间,令其自然转移。接蚜8 d后进行查蚜。查蚜方法为计数法。每行查蚜10株,以每株的平均蚜量定级,共分5级(表 1)。每品种抗性鉴定重复3次F1代采用群体评级,F2代采用单株评级。

|
| 图 1 抗蚜辣椒品种‘猪大肠’和感蚜辣椒品种‘大羊角椒’正反杂交 |
从‘猪大肠’和‘大羊角椒’正反杂交的F2代群体中各选取10株经过抗性鉴定的抗、感蚜辣椒用于辣椒POD和PPO酶活分析。POD和PPO的酶活分析方法参照Xiao等[12]的方法进行。
1.2.3 数据分析采用SPSS软件进行数据分析,显著性差异分析采用One-Way ANOVA-Fisher中的LSD方法,所有数据均为3个生物学重复的平均值,显著性检测水平为α=0.05。
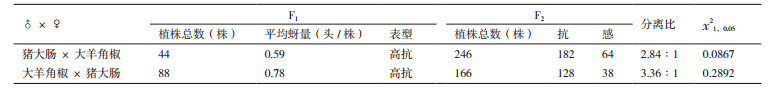
2 结果 2.1 辣椒抗蚜性的遗传分析抗蚜性鉴定结果表明,‘猪大肠’和‘大羊角椒’正反杂交后代F1表型均没有出现抗、感性分离,平均每株蚜量分别为0.59头和0.78头,表现为高抗(HR)。F1自交所得F2表型出现抗、感性分离,抗蚜与感蚜植株分离比经χ2测验符合3:1分离规律(表 2)。上述结果说明,辣椒抗蚜性由单显性核基因控制,且能稳定遗传。此外,图 2结果表明,感蚜辣椒植株在接种桃蚜8 d后症状明显,而抗蚜辣椒植株只表现出轻微的桃蚜为害症状。

|
| 图 2 典型的抗、感蚜辣椒植株接种前后症状对比图 |
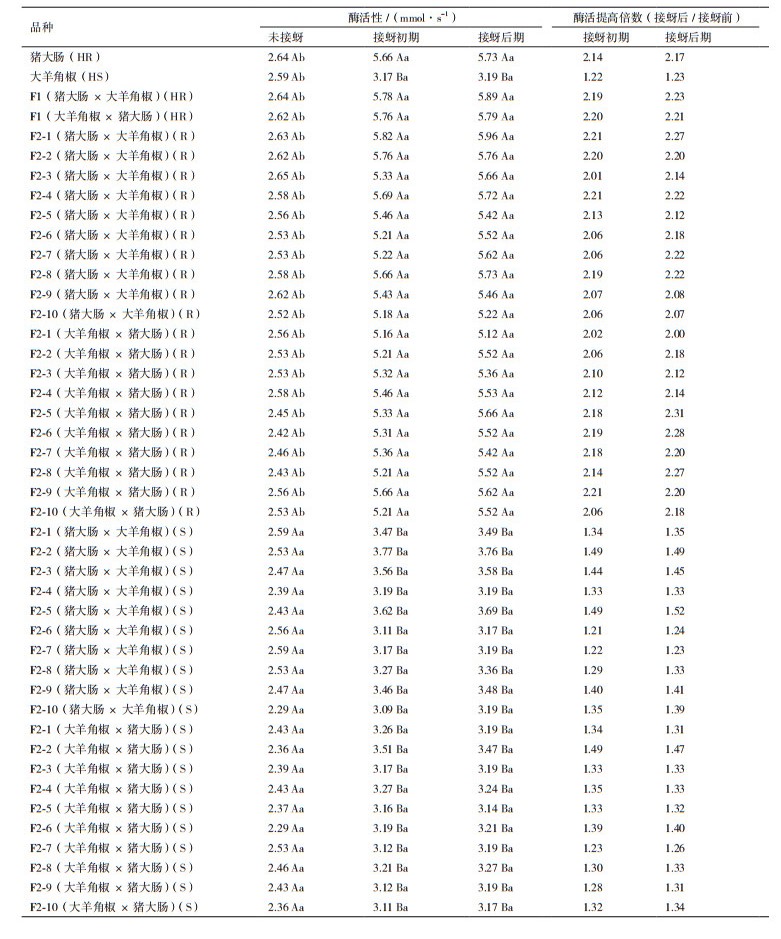
表 3结果表明,无论对于辣椒亲本及其F1、F2代群体,抗蚜品种在接蚜初期和后期植株叶组织内PPO活性并无显著差异,但与未接蚜相比均显著升高(P < 0.05),升高倍数在2.01倍-3.07倍之间,而感蚜品种PPO活性在接蚜前后均无显著差异(P < 0.05)。此外,接蚜前抗、感蚜辣椒品种PPO活性也无显著差异,而抗蚜品种接蚜初期、后期叶组织内PPO活性也分别显著高于感蚜品种(P < 0.05)。

表 4结果表明,无论对于辣椒亲本及其F1、F2代群体,抗蚜品种在接蚜初期和后期植株叶组织内POD活性并无显著差异,但与未接蚜相比均显著升高(P < 0.05),升高倍数在2.00倍-2.31倍之间,而感蚜品种POD活性在接蚜前后均无显著差异(P < 0.05)。此外,接蚜前抗、感蚜辣椒品种PPO活性也无显著差异,而抗蚜品种接蚜初期、后期叶组织内PPO活性均分别显著高于感蚜品种(P < 0.05)。
作物与有害生物的互作是一个长期进化中互相影响的过程,当我们利用植物在长期进化中积累的抗性来培育抗虫品种时,抗性的长期稳定遗传是我们首先要考虑的问题[13]。单基因抗性遗传简单,便于操作,而多基因抗性由于随着代数的增加而逐渐分散,使抗性削弱。对春小麦[14]、桃树[13]、高粱[15]、苜蓿[16]等作物的抗蚜性遗传分析表明,其抗蚜性均为质量性状遗传,其抗性各由一对显性单基因控制。本研究发现F1自交所得F2表型出现抗感性分离,抗虫与感虫植株分离比经χ2测验符合3:1分离规律,表明辣椒抗蚜性由单显性核基因控制,且能稳定遗传。在大羊角椒×猪大肠的F2代分离群体中,感虫植株的数量比预期3:1分离比率略高,根据Zhang等[17]的解释,此现象可能是由于感虫品种中存在一个修饰基因,改变了显性基因的抗性。
PPO和POD参与植物的防御反应,其活性显著升高可有效提高植物抗虫性水平[18-19]。PPO和POD不仅可通过清除活性氧而抵御昆虫对植物的侵害,还可作为次生代谢途径中的重要酶,调控和影响寄主植物次生代谢物质的产生,从而增强植物的抗虫性[20-21]。研究发现,虫害胁迫的茶树叶片PPO和POD等含量升高,活性增强,对昆虫及其诱发的一系列病害有防御作用[19, 22]。麦长管蚜为害可诱导抗性小麦叶组织中PPO和POD活性显著升高,而敏感种质中却无显著变化[18]。桃蚜为害可诱导甘蓝体内PPO和POD活性的升高,且随桃蚜为害时间的延长而增加[23]。蚜虫为害后,抗性高梁种质体内PPO和POD活性比感蚜前显著升高,并与蚜虫的诱导相关[24]。蚜虫取食可诱导棉花体内的PPO和POD活性升高,其升高速度及峰值高低与棉花抗蚜性呈正相关关系[25]。本研究针对PPO和POD酶活分析结果与前人相似,抗性辣椒种质受蚜虫为害初期叶组织PPO和POD活性不仅显著高于为害前水平,而且显著高于感蚜品种,并且可以将酶活维持在一个稳定的高水平至接蚜后期,表明辣椒PPO和POD活性增加幅度与辣椒抗蚜性具有显著相关性。
4 结论针对辣椒抗蚜品种‘猪大肠’的抗蚜性遗传分析结果表明,辣椒抗蚜性由显性单基因控制,能够稳定遗传。PPO和POD酶活性受桃蚜为害后显著升高与抗蚜辣椒品种的抗性显著相关。
| [1] |
汤秋玲, 马康生, 高希武. 蔬菜蚜虫抗药性现状及抗性治理策略[J]. 植物保护, 2016, 42(6): 11-20. |
| [2] |
陈青. 几种生化物质与辣椒抗蚜性的相关性[J]. 园艺学报, 2002, 29(6): 533-536. |
| [3] |
任广伟, 王凤龙, 彭世阳. 马铃薯Y病毒与介体蚜虫传毒的关系研究进展[J]. 烟草科技, 2006(10): 56-61. DOI:10.3969/j.issn.1002-0861.2006.10.015 |
| [4] |
王佳, 王亚峰, 蒲颇, 陈媛, 刘映红. 烟草感染两种病毒对烟蚜种群增长、寄主选择与传毒的影响[J]. 西南大学学报:自然科学版, 2017, 39(3): 23-27. |
| [5] |
钟秀明, 武雪萍. 我国农田污染与农产品质量安全现状、问题及对策[J]. 中国农业资源与区划, 2007, 28(5): 27-32. |
| [6] |
赵善欢, 张兴. 植物自身控制害虫的作用[J]. 华南农业大学学报, 1985, 6(2): 79-89. |
| [7] |
Sarath BB, Pandravada SR, Rdvj PR, et al. Global sources of pepper genetic resources against arthropods, nematodes and pathogens[J]. Crop Protection, 2011, 30(4): 389-400. DOI:10.1016/j.cropro.2010.12.011 |
| [8] |
Frantz JD, Gardner J, Hoffmann MP, et al. Greenhouse screening of Capsicum accessions for resistance to green peach aphid(Myzus persicae)[J]. HortScience, 2004, 39(6): 1332-1335. |
| [9] |
Bosland PW, Ellington JJ. Comparison of Capsicum annuum and C. pubescens for antixenosis as a means of aphid resistance[J]. HortScience, 1996, 6: 1017-1018. |
| [10] |
孙蕾, 程须珍, 王素华, 等. 栽培绿豆V2709抗豆象特性遗传分析[J]. 植物遗传资源学报, 2007, 8(2): 209-212. |
| [11] |
陈青. 辣椒抗蚜性的生化基础及其RAPD分析[D]. 海口: 海南大学, 2002.
|
| [12] |
Liang X, Chen Q, Lu H, et al. Increased activities of peroxidase and polyphenol oxidase enhance cassava resistance to Tetranychus urticae[J]. Exp Appl Acarol, 2017, 71(3): 195-209. DOI:10.1007/s10493-017-0125-y |
| [13] |
牛良, 鲁振华, 曾文芳, 等. '粉寿星'对桃绿蚜抗性的遗传分析[J]. 果树学报, 2016(5): 578-584. |
| [14] |
胡秉芬, 马小乐, 师桂英, 等. 春小麦抗蚜品系'J-31'和'J-48'的抗性遗传分析[J]. 甘肃农大学报, 2009, 44(3): 58-63. |
| [15] |
常金华, 夏雪岩, 张丽, 等. 高梁抗蚜基因的遗传分析和SSR标记定位[J]. 草业学报, 2006, 15(2): 113-118. |
| [16] |
郭素敏. 蒺藜苜蓿抗蚜虫的遗传分析[D]. 南京: 南京林业大学, 2011.
|
| [17] |
Zhang XP, Rhodes B. Inheritance of resistance to races 0, 1, and 2 of Fusarium oxysporum f. sp. niveum in watermelon(Citrullus sp. PI 296341)[J]. Report-Cucurbit Genetics Cooperative, 1993, 21(6): 121-132. |
| [18] |
Ying H, Yu W, Bi JL, et al. Constitutive and induced activities of defense-related enzymes in aphid-resistant and aphid-susceptible cultivars of wheat[J]. J Chemi Ecol, 2009, 35(2): 176-183. DOI:10.1007/s10886-009-9589-5 |
| [19] |
Yang ZW, Duan XN, Jin S, et al. Regurgitant derived from the tea geometrid Ectropis obliqua suppresses wound-Induced polyphenol oxidases activity in tea plants[J]. J Chem Ecol, 2013, 39(6): 744-751. DOI:10.1007/s10886-013-0296-x |
| [20] |
Van Damme EJM. Induced Plant Resistance to Herbivory[M]. 2nd ed. New York: Springer, 2008.
|
| [21] |
Zhurov V, Navarro M, Bruinsma KA, et al. Reciprocal responses in the interaction between Arabidopsis and the cell-content-feeding chelicerate herbivore spider mite[J]. Plant Physiology, 2014, 164(1): 384-392. DOI:10.1104/pp.113.231555 |
| [22] |
王丹, 陈亮. 茶树对茶尺蠖抗性机制研究[J]. 茶叶科学, 2014(6): 541-547. |
| [23] |
张春妮. 甘蓝苗期对桃蚜Myzus persicae(Sulzer)抗性的生化机制研究[D]. 杨陵: 西北农林科技大学, 2005. http://www.lunwentianxia.com/product.sf.3497003.1/
|
| [24] |
张丽, 常金华, 罗耀武. 不同高粱基因型感蚜虫前后POD、PPO、PAL酶活性变化分析[J]. 中国农学通报, 2005, 21(7): 40-48. |
| [25] |
李润植, 毛雪, 李彩霞, 等. 棉花诱导抗蚜性与次生代谢相关酶活性的关系[J]. 山西农业大学学报:自然科学版, 1998(2): 165-168. |