




2. 中国农业科学院生物技术研究所 农业部转基因安全评价重点实验室,北京 100081
2. MOA Key Laboratory on Safety Assessment(Molecular)of Agri-GMO, Chinese Academy of Agricultural Sciences, Beijing 100081
水稻(Oryza sativa L.)是世界上最重要的粮食作物之一,为亚洲半数以上人口的主食,在人类经济、文化发展过程中起到了非常重要的作用。水稻种植过程中容易受到诸多病、虫及草害等危害。为保证种植作物稳产和高产的前提,提高作物品质,增加经济收益,人们试图将生物技术与作物遗传改良相结合,通过提高水稻抗性的方式来提高水稻产量。近年来,研究人员培育出一大批品质优良,遗传稳定的转基因作物,在给农业带来巨大的社会效益和经济效益的同时,其生态安全性和食品安全性也备受关注[1],这也是目前转基因水稻还未商业化的主要因素。因此在田间条件下进行转基因水稻生态安全风险评估已成为目前研究的重点,而研究转基因水稻的基因飘流是其生态安全评估的主要内容之一[2-3]。
基因飘流是指同种或异种有性可交配物种之间通过风媒或虫媒、风媒加虫媒及供体的花粉扩散到受体植物的柱头上授粉、受精结实的过程,简称由花粉扩散介导基因飘流[4]。不同类型的水稻花粉导致基因飘流差异较大,近10多年来,国内外以转基因水稻作为花粉供体,以栽培稻、普通野生稻、不育系等作为受体,采用长方形或同心圆田间试验设计进行大量试验,阐明了不同类型的水稻基因飘流的基本规律,并建立了可预测水稻转基因飘流的高斯烟羽模型[5]。研究结果表明,水稻作为自花授粉植物,天然异交结实率低,一般小于1%[6-7],转基因水稻与非转基因水稻相邻种植时,供体向不育系的基因飘流率最高,为1.556%(培矮64S)-80.100%(中9A),平均为46.25%。向普通栽培稻的最大基因飘流率 < 1%,平均为0.33%[8]。向普通野生稻的基因飘流率介于不育系和常规稻之间,为11.24%-18.00%[4]。同时转基因水稻也可以向红稻和杂草稻进行基因飘流,向杂草稻和红稻的基因飘流率小于1%[9-11]。
栽培稻在遗传上可分为两个亚种,即籼亚种和粳亚种,由于两个亚种的开花习性和异交结实特性有差异,因而作为花粉源的不同亚种基因飘流存在差异。基因飘流影响因子包括水稻花粉扩散能力或竞争力、受体的异交结实能力等。因此,研究基因飘流并准确建立基因飘流模型需要分析水稻花粉竞争力。水稻花粉竞争力包括数量竞争力与遗传竞争力,数量竞争力是当两个供体的遗传背景相同时,因花粉数量不同引起的对基因飘流率的数值影响,遗传竞争力是当两个供体的花粉数量相同时,因供/受体的遗传亲和力不同所造成的对基因飘流率的数值影响。研究表明,在同等数量条件下,粳型转基因水稻B2与籼型转基因水稻B9向不育系博A的基因飘流存在一定差异,表现为在同一距离上两者的基因飘流频率差异极显著(P < 0.01)。当受体为不育系时,无论花粉源是籼型还是粳型转基因水稻,在不同距离点的基因飘流频率均为粳型 > 籼型[12]。此外,在杂交水稻的制种实践中,父母本行比是影响杂交水稻制种产量的重要因素,随行比数量增加,供体花粉量增加,制种产量增加[13-14],这意味着花粉源数量或源强大小影响受体的异交结实率。因此,在研究基因飘流时,除了要考虑花粉源的籼、粳类型之外,还需要研究花粉供体数量或源强大小对基因飘流的影响。
以往的研究大多只研究了一种花粉供体向不同受体的基因飘流率及差异,即使有个别研究不同水稻亚种类型的基因飘流差异的例子,也没有考虑两者之间的数量关系对基因飘流的共同影响。另一方面,过去的研究只考虑水稻供体与受体之间的数量竞争性,忽略了不同水稻供体之间对受体的竞争性,更没有考虑花粉源数量竞争性和遗传竞争性的相互作用对基因飘流的影响。鉴于此,为了研究籼稻与粳稻的遗传竞争性与花粉源数量配比对基因飘流的影响,我们设计籼、粳双供体在不同行比的条件下的基因飘流的试验,试图解析转基因水稻花粉源的在基因飘流中的数量竞争性和遗传竞争性。水稻的基因飘流率受到花粉竞争力的影响,它应该是改善水稻基因飘流模型精度的重要途径。但到目前为止,有关转基因水稻花粉的数量竞争和遗传竞争的研究尚未见报道。开展对转基因水稻花粉竞争力的研究,对进一步探讨水稻基因飘流变化规律、认识和优化水稻转基因飘流模型具有重要意义。
1 材料与方法 1.1 材料花粉供体:转基因水稻为粳稻B2和籼稻Bar9311(代号B9); 常规稻为粳稻秀水63(代号XS63)和籼稻9311。花粉受体为不育系博A。
B9为含bar基因抗除草剂的转基因籼稻品系(Oryza sativa L. cv. Indica),该品系是由转基因抗除草剂恢复系Bar68-1与9311经过杂交并多次回交获得的稳定株系材料[15]。
B2为含bar基因的抗BastaR转基因品种(Oryza saliva L.cv. Japonica),由中国水稻研究所利用基因枪法将bar基因导入到常规粳稻京引119中获得转bar基因粳稻植株T京引119[16],然后将其与常规粳稻秀水63杂交获得B2植株[17]。
1.2 方法 1.2.1 田间布局本试验于2016年12月于海南省乐东县海南大学尖峰基地进行,试验地位于东经108°46'12″,北纬18°38'39″,试验区域平坦开阔,水稻种植的行向和盛行风向垂直。水稻播种后采用湿润育秧,按要求人工移栽,移栽规格20 cm×20 cm。
为了研究花粉竞争性,本试验将转基因水稻和常规稻4种按照“转基因+非转基因”双供体组合配比设置花粉源,检测其基因飘流。转基因与非转基因的花粉源配比可以通过检测确定受体(不育系)接受的花粉来自哪一个供体。花粉源组合包括B9(籼,转基因)+9311(籼)、B9(籼)+XS63(粳)、B2(粳,转基因)+9311(籼)、B2(粳)+XS63(粳)配对,每个花粉源组合均以1:9、2:8、3:7、4:6、5:5、6:4、7:3、8:2、9:1的行比种植在相互隔离的小区中,田间布局为4 m×4 m的小区,共计有36个小区,供体与受体(不育系博A)相邻种植,不同的小区用2 m高隔离布相互隔离(图 1)。根据不同的花粉供体、按照不同的行比来模拟小区中花粉的量的差异,通过不同小区转基因与常规稻种植行比的变化和双供体类型的变化,来模拟不同花粉量和籼、粳两种亚种类型对基因飘流的影响。

|
| 图 1 花粉竞争对基因飘流影响试验田间布局 左图双供体试验不同比例的小区试验图,右图(上图)为双供体类型(下图)为小区内部栽插布局图,黑色边框表示隔离布阻挡 |
根据预备试验推测供体、受体的生育期,确保花期相遇。花期重叠天数至少5-7 d,并记录水稻生长的播插期、始穗期、齐穗期、始花期、终花期及成熟收获期等各个时期。花粉供体B9、9311、XS63、B2和受体的花期相遇良好,各材料花期重叠日数在10 d以上。收获时,单个小区样品混收,分成3份,作为转基因检测的3次重复。
1.2.3 田间样品检测基因飘流试验结束后,收获全部种子并进行下一步田间检测。样本种子种植后在3叶期开始喷施BastaR除草剂,每周一次,共喷施3次,3次浓度为依次为500 mg/L、500 mg/L和800 mg/L。同时,种植供体和受体材料作为阳性和阴性对照。喷施后存活的种子采用PCR检测,排除假阳性。按照Jia等[8]的统计公式,计算基因飘流频率(%):

按照图 1种植的供、受体材料,抽穗开花后受体完成异交结实,成熟后收获种子,另选秧地播种,待水稻秧苗长至3叶期,通过喷施3次除草剂BastaR,可直观辨别检测转基因后代。主要表现为绿色成活抗性苗是转基因后代,枯死苗为常规稻杂交后代。为避免抗BastaR苗的假阳性,随机选取绿色抗性苗,采用CTAB法提取DNA进行分子生物学检测[18]。PCR结果显示绿色存活苗均含有bar基因,如图 2为bar基因条带,1-22均有表达,排除了假阳性。

|
| 图 2 转基因水稻基因飘流后代抗除草剂苗PCR结果 M:Marker 500;“+”阳性对照; “-”阴性对照 |
为研究不同行比下基因飘流的数量竞争性,试验设计不同的行比比例,以不同行比种植在相互隔离的小区中,各小区单独隔离,保证各小区花粉传播的独立性。转基因与非转基因种植呈比例变化。理论上讲,相同亚种姊妹系的花粉竞争力相同,因此试验设计B9+9311与B2+XS63相同亚种姊妹系作为数量竞争性的双供体,以此研究不同行比竞争导致的基因飘流的差异。通过趋势拟合发现基因飘流率与行比的关系可用公式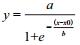
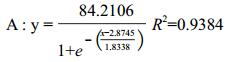 (1)
(1)
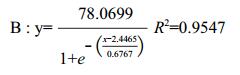 (2)
(2)

|
| 图 3 转基因与非转基因水稻双供体行比与基因飘流率关系 横坐标为小区中转基因与非转基因种植行比1:9,2:8,3:7,4:6,5:5,6:4,7:3,8:2,9:1 |
因此,基因飘流率与供体行比成S曲线关系,基因飘流随供体行比增加,在转基因行比数量较小的条件下,基因飘流率变化较小,随转基因水稻行比数量的增加,基因飘流率显著增加; 在行比数量达到一定值时,基因飘流率增加不明显。
转基因与非转基因行数相同,籼、籼配比(B9:9311)基因飘流比例接近1:1;粳、粳配比(B2:XS63)基因飘流的比例大于1:1。即籼型花粉源同亚种类型的行比数量相同,两个供体之间基因飘流的竞争力差异不显著。这从另一侧面印证了相同籼亚种姊妹系花粉供体的竞争性基本相同。
2.3 籼、粳亚种花粉供体基因飘流的竞争性不同类型的花粉供体基因飘流存在差异,通过不同花粉供体的亚种类型配置和行比配置,研究由不同供体导致的基因飘流差异,探究拟合水稻双供体数量竞争性及遗传竞争性曲线模型,分析基因飘流竞争性。表 1是不同籼、粳两个亚种双供体在不同行比配置下的基因飘流率的平均数差异。由表 1可知,组合A和D差异极显著(P < 0.01),飘流率均值A(58.277 8) > D(19.890 0),即(B9+9311)组合与(B2+9311)组合在相同籼型供体9311竞争的条件下,B9飘流率与B2飘流率存在显著差异,即B9的竞争性大于B2;组合B和C差异显著(P < 0.05),飘流率均值B(61.390 0) > C(40.434 4),即(B9+XS63)组合与(B2+XS63)组合在相同粳稻供体XS63的竞争条件下,B9飘流率与B2飘流率存在显著差异,B9的竞争性大于B2。可见,转基因籼稻的竞争性大于转基因粳稻。
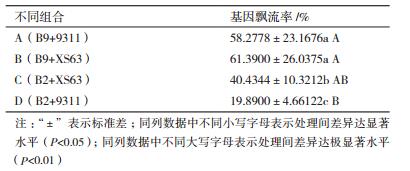
组合C和D差异显著(P < 0.05),飘流率均值C(40.434 4) > D(19.890 0)。(B2+XS63)组合与(B2+9311)组合有相同的粳型转基因供体B2,XS63和9311提供了各自小区的常规花粉竞争,C > D说明D区中竞争花粉的能力更强,即9311的竞争性大于XS63。因此常规籼稻竞争性大于常规粳稻,即籼稻花粉的竞争性大于粳稻。
3 讨论本试验结果表明,水稻花粉的竞争性包括数量竞争性和遗传竞争性。当两个供体的遗传背景相同时,因水稻花粉量不同而引起的基因飘流的差异可称之为数量竞争性。当两个供体的遗传背景不相同时,因异交结实不同引起的基因飘流的差异可称为遗传竞争性。同种类型的水稻不同种植比例下的基因飘流率随着转基因行比的增加而增大。籼、粳双供体因两个供体之间遗传背景不同,其基因飘流率的变化既存在遗传竞争性的差异,又存在数量竞争性的差异,不同种植比例下的基因飘流率应为两种竞争性共同作用。前人只研究同一供体对不同受体的基因飘流,没有研究不同供体的基因飘流竞争性。本试验弥补了前人研究的不足。
3.1 转基因水稻基因飘流的数量竞争性在对水稻进行花粉竞争性研究时,在数量竞争性作用下,水稻相同亚种在不同行比配制下的基因飘流率呈S曲线关系。在转基因行比数量较小的条件下,转基因颖花数量少,受体接受花粉较少,因此基因飘流率变化较小; 随转基因水稻行比数量的增加,颖花数量增加,受体接收花粉的数量显著增加,基因飘流率显著增加; 在行比数量达到一定值时,颖花数量接近饱和,基因飘流率增加不明显。同时,在杂交水稻的制种实践中,父母本行比是影响杂交水稻制种产量的重要因素,随行比数量增加,供体花粉量增加,制种产量增加[13-14]。因此,供体颖花数量是影响基因飘流数量竞争性的主要因素。籼、籼配比的基因飘流比例与粳、粳配比的基因飘流的比例都接近1:1,在相同行比条件下,基因飘流率相同。即同种亚种类型配比,基因飘流的竞争差异不明显,可视为具有同等基因飘流能力。
3.2 转基因水稻基因飘流遗传竞争性在相同行比配置条件下,比较籼、粳两种亚种的花粉竞争,结果表明籼稻与粳稻花粉竞争存在显著差异,籼稻的竞争性大于粳稻。不同供体相同栽插行比的条件下,籼稻的竞争性大于粳稻。分析其原因,认为籼稻单株分蘖数量优于粳稻,籼稻的优势主要是根系发达、分蘖能力强、穗大花粉量多[19],导致籼稻单位面积的花粉数量大于粳稻,同等行比情况下籼稻花粉数量占据优势。何美丹等[20]比较了两组平行试验中的籼、粳型转基因水稻基因飘流差异,分别以籼型转基因水稻B9和粳型转基因水稻B2为花粉供体,籼型不育系博A为受体材料,受体中同期种植常规籼稻特籼占25作为竞争花粉源,在同等自然条件下研究籼、粳型转基因水稻向不育系和常规稻的基因飘流频率。结果表明,当受体为不育系时,无论花粉源是籼型还是粳型转基因水稻,在不同距离点的基因飘流频率均为粳型大于籼型。10 m试验区内籼型转基因水稻向不育系各点平均基因飘流频率为1.325%-23.948%,粳型为3.906%-45.934%[12, 20]。但我们注意到,该试验在相邻距离种植了与受体不育系BoA相同花期的籼型竞争花粉源特籼占25,且不育系BoA与常规稻特籼占25按插植行比3:1相间种植,试验中籼型花粉源的竞争性较大。另一方面,同为籼稻的花粉供体和竞争花粉源植株每日开花时间(花时)基本相同,籼稻比粳稻花时早而且持续时间较长,而籼、粳花时相遇时间较短[21],这更是增加了对籼稻花粉源的竞争性。而在该试验中粳稻转基因花粉源并没有来自同类型常规粳稻花粉的竞争,因而该试验中两组基因飘流结果相比较,其结论是在不同距离的取样点粳稻的基因飘流频率均大于籼稻。因此,在籼型花粉源竞争能力强大的情况下,不同距离点上粳稻基因飘流频率大于籼稻是可以理解的。可见,在研究转基因飘流的遗传竞争过程中,数量竞争的影响是很大的,也许在一定条件下水稻的数量竞争占主导作用。
此外,研究遗传竞争性需要基于籼、粳两种亚种小区中的颖花数量完全相同的情况,单纯计算两个亚种的行比来确定遗传竞争性,似乎包括了籼粳两种花粉供体植株群体数量不同而导致的花粉数量竞争,这种竞争性是数量竞争与遗传竞争共同作用的综合结果,且在一定条件下水稻的数量竞争占主导作用。本研究的结论“在相同行比配置条件下,籼稻的竞争性大于粳稻”应该是数量竞争与遗传竞争共同作用。另外,不同的研究结果比较还要考虑试验季节、气候类型和花期相遇程度等,有关结论有待进一步研究探讨。
4 结论水稻相同亚种的籼-籼与粳-粳双供体组合的基因飘流频率受数量竞争性的影响; 籼-粳不同亚种双供体的基因飘流频率,受花粉源的数量竞争性和遗传竞争性的共同影响。相同亚种花粉源组合的数量竞争性表现为基因飘流率与转基因花粉供体行比呈S型曲线的关系; 籼-粳花粉源组合在数量竞争与遗传竞争共同作用下,籼稻基因飘流率大于粳稻,说明籼稻花粉竞争性强于粳稻。花粉数量竞争性和遗传竞争性共同影响水稻的基因飘流。
致谢: 本研究所使用的水稻材料B2由中国水稻研究所钱前博士提供,Bar9311由中国科学院亚热带农业生态研究所肖国樱博士提供,谨致谢忱。
| [1] |
张启发. 大力发展转基因作物[J]. 华中农业大学学报:社会科学版, 2010, 85(1): 1-6. |
| [2] |
孙国庆, 金芜军, 宛煜嵩, 等. 中国转基因水稻的研究进展及产业化问题分析[J]. 生物技术通报, 2010(12): 1-6. |
| [3] |
Ellstrand NC. Current knowledge of gene flow in plants:implications for transgene flow[J]. Philosophical Transactions of the Royal Society of London, 2003, 358(1434): 1163. DOI:10.1098/rstb.2003.1299 |
| [4] |
裴新梧, 袁潜华, 胡凝, 等. 水稻转基因飘流[M]. 北京: 科学出版社, 2016.
|
| [5] |
胡凝, 陈万隆, 刘寿东, 等. 水稻花粉扩散的模拟研究[J]. 生态学报, 2010, 30(14): 3665-3671. |
| [6] |
姜大刚, 梁奕涛, 黄靖, 等. 转基因水稻外源基因的漂移研究[J]. 生物技术通报, 2010(6): 95-99. |
| [7] |
Oka HI. Origin of Cultivated Rice[M]. Elsevier/churchill Living-stone, 1988: 962.
|
| [8] |
Jia S, Wang F, Shi L, et al. Transgene flow to hybrid rice and its male-sterile lines[J]. Transgenic Research, 2007, 16(4): 491-501. DOI:10.1007/s11248-006-9037-z |
| [9] |
Messeguer J, Marfà V, Català MM, et al. A field study of pollen-mediated gene flow from mediterranean Gm rice to conventional rice and the red rice weed[J]. Molecular Breeding, 2004, 13(1): 103-112. DOI:10.1023/B:MOLB.0000012285.39859.9d |
| [10] |
Lentini Z, Debouck D, Espinoza AM, et al. Gene flow analysis into wild/weedy relatives from crops with center origin/diversity in tropical america[M]. Saskatoon: International Society for Biosafety Research, 2006: 152-156.
|
| [11] |
程林, 韩飞, 袁潜华. 转基因稻向栽培稻及其稻属植物的基因漂流研究进展[J]. 贵州科学, 2007, 25(4): 42-46. DOI:10.3969/j.issn.1003-6563.2007.04.008 |
| [12] |
何美丹, 徐立新, 尹昭坤, 等. 籼、粳型转基因水稻花粉源基因飘流差异研究[J]. 南方农业学报, 2012, 43(5): 559-565. DOI:10.3969/j:issn.2095-1191.2012.05.559 |
| [13] |
黄光福.父母本行比和密度对杂交水稻制种产量形成的影响[D].长沙: 湖南农业大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10537-1017033787.htm
|
| [14] |
谢必武, 张凤龙, 晏承兴, 等. 川农优528机械化制种技术研究[J]. 种子, 2013, 050(6): 123-127, 134. DOI:10.3969/j.issn.1001-4705.2013.06.039 |
| [15] |
肖国樱, 唐俐, 袁定阳, 等. 转Bar基因抗除草剂两系杂交早稻恢复系Bar 68-1的培育研究[J]. 杂交水稻, 2007, 22(6): 57-61. DOI:10.3969/j.issn.1005-3956.2007.06.023 |
| [16] |
朱冰, 田文忠. 利用基因枪法获得可遗传的抗除草剂转基因水稻植株[J]. 中国农业科学, 1996, 29(6): 15-20. DOI:10.3321/j.issn:0578-1752.1996.06.003 |
| [17] |
华志华, 朱雪峰, 吴明国, 等. 水稻转基因整合模式中外源基因的遗传规律[J]. 作物学报, 2003, 29(1): 44-48. |
| [18] |
王关林, 方宏筠. 植物基因工程[M]. 北京: 科学出版社, 2009.
|
| [19] |
吴一才. 杂交粳稻分蘖的初步研究[J]. 辽宁农业科学, 1980(3): 13-17. |
| [20] |
何美丹.利用物理屏障控制转基因水稻基因飘流的研究[D].海口: 海南大学, 2012. http://cdmd.cnki.com.cn/Article/CDMD-10589-1015505882.htm
|
| [21] |
王开锡, 林庆泉, 杨聚宝. 籼、粳稻花时的分布及相遇情况的调查[J]. 杂交水稻, 1990(5): 19-20. |