




类固醇雌激素作为环境激素中雌激素效应最高一类物质[1],环境曝露会导致动物雄性繁殖能力下降、生殖系统发育异常,男性精子数量减少、质量下降,女性生理现象混乱,内分泌系统异常亢奋或抑制,神经内分泌功能紊乱,激素依赖性器官肿瘤发病率明显升高等[2, 3]。雌酮(Estrone,E1) 作为具有雌效应的天然类固醇雌激素之一,在极低的浓度水平即可使水生生物性腺功能异常,具有高毒性、难降解的特性,通过食物链在体内蓄积[4],进而威胁人类和动物的健康。类固醇雌激素主要由人与动物卵巢合成分泌,通过污水处理厂和禽畜养殖场两种主要途径进入环境[5, 6]。研究表明处于孕期的女性E1排泄量最多可达到1 194 µg/d[7]。国内外各种水体及污水厂出水中均检测到较高浓度的E1[8, 9]。2007年,中国天然类固醇雌激素的总排放量高达10.6 t[10]。E1被土壤微生物降解转化的同时,微生物也会受到E1的影响(如细胞形态结构改变、酶的产生及效用改变和DNA突变等),具体表现在种群的生长与消亡;不同种微生物对E1的敏感程度不同,土壤微生物群落作为混合性的微生物群,E1作用下表现为群落结构发生变化。土壤微生物群落结构的改变将直接影响该植被上农作物的生长状况,进而影响高级生态系统的健康。
本文以雌酮作为胁迫因子,以农田土壤作为实验材料,通过PCR-DGGE技术探讨不同雌酮浓度、不同培养时间下农田土壤微生物群落变化,旨在了解雌酮胁迫下农田土壤微生物群落动态变化特征。
1 材料与方法 1.1 材料供试土壤:农田土壤,采自沈阳农业大学水稻试验田。该土壤常年种植水稻,并未施用畜禽粪便或市政污水。主要理化性质如下:有机质含量23.6 g/kg,总氮1.54 g/kg,有效磷71.6 mg/kg,总磷1.14 g/kg,速效钾120 mg/kg,总钾16.7 g/kg,pH值7.62,类固醇雌激素未检出。
1.2 方法 1.2.1 实验设计本实验的主要培养装置为塑料盒,其长度、宽度和高度分别为490、350和160 mm。将15 kg的农田土壤装入塑料盒,500 mL不同浓度的雌酮溶液分别加入到各塑料盒中,最后使得各装置的终含量分别为500、1 000、1500和2 000 ng/kg,于室温培养。通过搅拌器使各溶液与农田土壤样品混合,以确保所有的农田土壤样品充分混合。每个塑料盒每次收集20 g再次充分混合的土壤样品,重复3次,曝露时间分别为0、5、10、15、20、25、30、35、40、45、50、55和60 d。将样品保存在-80℃用于分子生物学分析。
1.2.2 土壤中总DNA提取本实验使用E.Z.N.ATM Soil DNA Kit试剂盒(美国OMEGA公司)提取农田土壤样品中的总DNA,用1.0%的琼脂糖凝胶电泳检测其提取结果。总DNA保存于-20℃冰箱中。
1.2.3 PCR-DGGE分析PCR的扩增区域是细菌16S r DNA的V3可变区,选用引物对338F和518R。选用的上游引物P1(5'端连接GC夹)为GC-338F(5'-CGC CCG CCG CGC GCG GCG GGC GGG GCG GGG GCA CGG GGGG CCT ACG GGA GGC AGC AG-3')和下游引物P2为518R(5'-ATT ACC GCG GCT GCT GG-3')[11],直接对总DNA进行PCR扩增。PCR反应体系为:模板DNA 0.1 μL,引物P1和P2各0.75 μL,2×GoTaq buffer 25 μL,无核酸酶水23.4 μL。PCR反应条件:94℃预变性5 min;94℃变性1 min,65℃退火1 min(每个循环降低1℃),72℃延伸1 min,一共10个循环;然后94℃变性1 min,55℃退火1 min,72℃延伸1 min,35个循环;最后72℃延伸10 min。PCR扩增产物用2.0%的琼脂糖凝胶电泳进行验证。所有的PCR产物约240 bp,可进行DGGE分析。
DGGE实验按照Webster等[12]方法进行并适当修改。PCR产物分离使用DCodeTM通用突变检测系统(Bio-Rad,Dcode,USA.)进行DGGE电泳分析。DGGE电泳条件为8%聚丙烯酰胺凝胶,变性梯度为35%-65%。电泳电压为150 V,电泳温度为60℃,缓冲体系是1×TAE缓冲液,电泳时间为7.5 h。电泳后,将凝胶用EB(溴化乙锭)染色30 min并在紫外光下观察。为获得清晰的图像,将使用凝胶照片系统(GelDoc,Bio-Rad,USA.)进行拍照。
1.2.4 DGGE图谱分析使用Quantity one图像分析软件对DGGE图谱进行分析(4.0版本,Bio-Rad,USA.)。该软件能够识别同一个凝胶不同泳道上占据相同位置的条带。两个样本的相似性可以通过戴斯系数(Cs)条带模式进行估测(公式1):
 (1)
(1)
其中,j是两条DGGE泳道中共有的条带数,a和b分别是两条DGGE泳道各自的条带数量。用非加权平均法(UPGMA)检验农田土壤样品的相似性。
基于DGGE条带数据而进行的香农-威纳指数(H'),丰富度(S)和均匀度(E)的计算比较了细菌多样性的变化。H'计算公式如下(公式2)
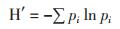 (2)
(2)
pi是指i条带出现在一条泳道中的概率。S是一条凝胶泳道的总条带数,E计算方法为H'/ lnS。
2 结果 2.1 雌酮对微生物群落结构的影响雌酮浓度为500、1 000、1 500、2 000 ng/kg时各个不同的时间段的细菌的16S rDNA基因的PCR产物DGGE电泳图谱如图 1,曝露时间分别为0、5、10、15、20、25、30、35、40、45、50、55和60 d。在分离图谱中,每个可以区分的条带都代表一个独立的细菌种群[13]。E1对农田土壤微生物群落结构变化产生影响。例如,条带1与条带4只存在于E1浓度为500和1 000 ng/kg的土壤样本中,条带6只存在于1 500与2 000 ng/kg浓度下,且颜色均是从无到有并逐渐变亮;而条带5在4个浓度下均存在。条带2与3仅存在于E1浓度为500 ng/kg下农田土样中,区别在于条带3在曝露前期就出现,条带2是从曝露中期出现。条带7仅出现在E1浓度为2 000 ng/kg的曝露前期,急速出现至鼎盛后又迅速消失。条带8出现于E1浓度为2 000 ng/kg的农田土样曝露中后期。

|
| 图 1 农田土壤在不同雌酮浓度不同暴露时间细菌16S rDNA的DGGE分析 A:雌酮浓度500 ng/kg;B:雌酮浓度1 000 ng/kg;C:雌酮浓度1500 ng/kg;D:雌酮浓度为2 000 ng/kg |
图 2是不同雌酮浓度下不同曝露时间的农田土壤样本与原始土壤样本的戴斯系数。与初始农田土壤样品相比,在第一个5 d内,由于刚加入雌酮,土壤微生物群落原始平衡被破坏,E1浓度为500 ng/kg的土壤样本的Cs平均下降了59.7%,E1浓度为1 000 ng/kg的土壤样本的Cs平均下降了17.1%,E1浓度为15 00 ng/kg的土壤样本的Cs平均下降了32.8%,E1浓度为2 000 ng/kg的土壤样本的Cs平均下降了62.8%。E1浓度为1 500 ng/kg的农田土壤的Cs直降到第10 d才暂时停止下降趋势,降低了55%。曝露时间为60 d时,E1浓度500 ng/kg农田土壤样品的Cs与初始样品只有30.6%的相似性;曝露时间为25 d时,E1浓度2 000 ng/kg农田土样的Cs仅为21.3%。E1浓度为500 ng/kg的农田土壤在第一个5 d之后,随着曝露时间的增加Cs波动平缓。E1浓度为1 000 ng/kg的农田土壤在曝露前期Cs的变化较缓和,第30 d之后Cs随着曝露时间的增加而剧烈变化。E1浓度为1 500 ng/kg的农田土壤样本在曝露时间内Cs始终都在剧烈波动。E1浓度为2 000 ng/kg的农田土壤的Cs在加入E1的前期剧烈波动,从第25 d到第40 d时波动相对平缓,第40 d后又出现巨大波动。

|
| 图 2 不同雌酮浓度不同暴露时间下农田土样微生物群落的戴斯系数 |
不同雌酮浓度的农田土壤样品,其系统进化树图谱有着显著差异(图 3)。在E1刺激的农田土壤样品中微生物群落结构表现出很大程度的变化,特别是E1浓度为2 000 ng/kg的农田土壤样品。结果表明,E1浓度越高,对微生物种群的影响越深,种群的亲缘关系也就越复杂。E1浓度为500 ng/kg、1 000 ng/kg、1 500 ng/kg、2 000 ng/kg时,曝露内所有实验土壤的遗传相似性分别为37%、71%、47%和39%。

|
| 图 3 不同雌酮浓度不同暴露时间农田土样的系统发育进化树 A:雌酮浓度为500 ng/kg;B:雌酮浓度为1000 ng/kg;C:雌酮浓度为1500 ng/kg;D:雌酮浓度为2000 ng/kg |
多样性指数能够有效地表征微生物群落结构的多样性。香农-维纳指数越高,微生物群落的多样性越大。除香农-维纳指数外,丰富度和均匀度也是多样性指数表征的重要参数:物种的总数即物种丰富度(S),不同物种个体数量的分布即物种均匀度(E)。
雌酮污染农田土壤样本中微生物群落的多样性指数如表 1所示。在曝露前,原农田土壤的H'在3.581-3.582之间,S均为53,E在0.908-0.909之间,说明在添加污染物前,农田土壤系统具有稳定的微生物群落结构和活性,同时具有基本的同质性。横向对比不同E1浓度发现,浓度为500 ng/kg和1 500 ng/kg的农田土壤中微生物群落多样性指数明显大于另外两组。E1浓度为500 ng/kg时农田土壤H'、S、E在曝露期间明显高于其余E1浓度曝露下的H'、S、E。E1浓度为1 000 ng/kg的农田土壤的多样性随着曝露时间的增加总体呈下降趋势;而E1浓度为500 ng/kg、1 500 ng/kg和2 000 ng/kg的农田土壤的多样性随着时间的推移总体趋势在逐步增长,且浓度为2 000 ng/kg时其增长趋势明显高于浓度为500 ng/kg与1 500 ng/kg时的增长趋势。纵向对比曝露时间,E1浓度为500 ng/kg农田土壤在第30 d时H'、S、E均高于其余曝露时间,但总体波动都比较平缓。E1浓度为1 000 ng/kg的土样在第20 d时H'、S、E达到最高值,之后在缓慢下降。浓度为2 000 ng/kg时,第40 d时的H'、S、E均达到最大值,且该浓度下多样性总体波动程度高于浓度为500 ng/kg和1 000 ng/kg时。E1浓度为1 500 ng/kg的农田土壤在曝露时间内,当S达到最大时,H'、E并没有达到最大,相比与其他浓度的微生物群落,此浓度下微生物群落显然是个不稳定的体系。
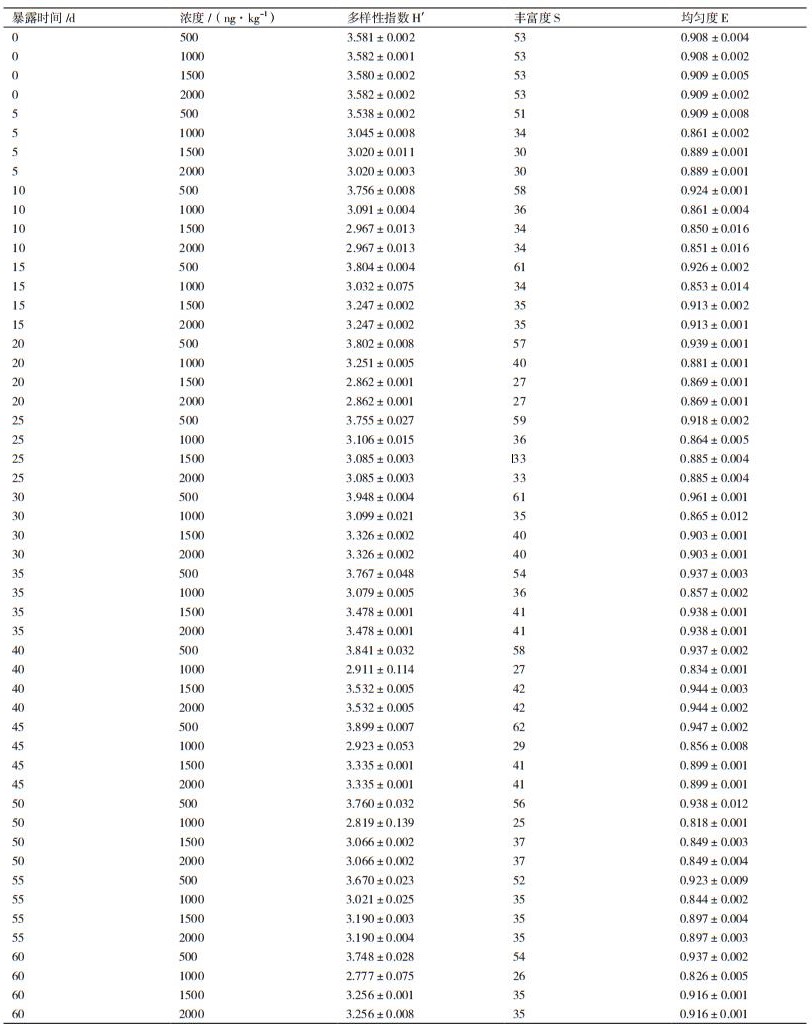
本实验相似性与多样性分析结果表明,在E1浓度为500 ng/kg至2 000 ng/kg之间,随着E1浓度的增大,其对微生物的影响增大,微生物生长受到抑制,多样性降低;可能因为原有的优势微生物因E1浓度的增大而逐步消亡(例如,条带2、3代表的种群),或者有可能已经出现新物种,只是新物种仍处于适应生长阶段,其在数量上并没有占据主要地位,故而在E1浓度为1 000 ng/kg的曝露后期微生物群落多样性都在不断下降;当超过某一浓度时,其对微生物的影响促使抗E1和可以有效降解E1的新型微生物(如条带6代表的种群)迅速适应、大量生长并逐渐占据主导地位,使得在E1浓度为1 500 ng/kg时,整个曝露期间内微生物多样性呈现平稳增长;继续加大浓度,E1为2 000 ng/kg时虽然在整个曝露阶段微生物多样性仍在缓慢持续增长,但是同1 500 ng/kg时增长趋势相比,此浓度下的增长趋势变缓,其原因有可能是原优势微生物受到一定的抑制作用进而消亡(例如,条带7所代表的种群),或者新式微生物出现并处于逐步适应阶段(例如,条带8所代表的种群)。
E1在土壤中的吸附量的多少会影响其被微生物降解转化的多少。刘建林[14]与吴乔龙[15]的研究表示,雌激素在土壤中吸附是自发放热过程,且在24 h内达到吸附平衡。而E1在土壤中的吸附作用受离子强度、pH、腐殖酸及其他共存污染物的共同作用。石月娥[16]发现E1在砂质壤土和粉砂性粘土中的吸附平衡时间延长约300%。
研究学者发现,E1与E2在生物降解过程中可以发生相互转化或单向转化,且在E1的降解过程中发现了E3[17]。同时E1在微生物酯化作用下也可形成结合态的E1-3G(G是葡萄糖苷酸)。类固醇雌激素之间的相互转化也会给微生物菌群多样性带来更为复杂的变化。张照韩[18]通过等摩尔比与等毒性单位复合的方式研究二元雌激素复合效应发现,除EE2和E1复合体系呈现拮抗作用外,其它复配体系均呈现协同效应,且复合作用效应的类型及强度与共存物质的种类及各种物质的比例有关。
王凌云[19]发现E1分子中C17-C13键级最低,因此E1可能会通过环开裂生成小分子羧酸等易降解物质。Ascenzo等[20]发现,即使经过最彻底的发酵E1仍有一定量的残留。目前对于环境介质(如土壤)中类固醇雌激素的代谢过程仍没有确定的途径,本实验可以进一步研究在该类农田土壤下E1的转化途径与代谢过程。不同种类固醇雌激素对微生物菌群的影响不同,这种情况与复合污染物类似。故针对单一污染物下分析微生物群落变化存在不确定的因素。因此,后续研究拟筛选有效降解类固醇雌激素的降解菌,并研究降解菌的降解特性。
4 结论PCR-DGGE指纹技术可以有效分析雌酮污染农田土壤样品中的微生物群落结构的动态变化。雌酮浓度和曝露时间的变化能够对微生物的活性和功能产生影响,进而造成微生物群落变化。
(1) 经戴斯系数与系统进化树图谱分析可知,雌酮浓度越高,对微生物种群的影响越深,种群的亲缘关系也就越复杂。(2) 经不同曝露时间与不同雌酮浓度微生物群落多样性指数对比分析可知,在一定浓度范围内,微生物群落多样性随着E1浓度的增加而减小,超过某一值时其多样性则会迅速增加,继续增大E1浓度,多样性增加趋势则逐渐变缓。(3) 雌酮可以转变为E2、E3以及在微生物酯化作用下形成结合态的E1-3G,这种相互转变也会给微生物菌群多样性带来更加复杂的变化。
| [1] | Khanal SK, Xie Bin, Thompson ML, et al. Fate, transport and biodegradation of natural estrogens in the environment and engineered systems[J]. Environmental Science and Technology, 2006, 40 (21): 6537–6546. DOI:10.1021/es0607739 |
| [2] | Treviño LS, Wang Q, et al. Hypothesis:Activation of rapid signaling by environmental estrogens and epigenetic reprogramming in breast cancer[J]. Reproductive Toxicology, 2015, 54 : 136–140. DOI:10.1016/j.reprotox.2014.12.014 |
| [3] | Tohyama S, Miyagawa S, Lange A, et al. Understanding the molecular basis for differences in responses of fish estrogen receptor subtypes to environmental estrogens[J]. Environmental Science & Technology, 2015, 49 (12): 7439. |
| [4] | 曹巧玲, 张俊明, 高志贤, 等. 环境内分泌干扰物研究的进展[J]. 中华预防医学杂志, 2007, 41(3): 224–226. |
| [5] | Kirk L A, Tyler C R, Lye C M, et al. Changes in estrogenic and androgenic activities at different stages of treatment in wastewater treatment works[J]. Environmental Toxicology & Chemistry, 2002, 21 (5): 972–979. |
| [6] | Lintelmann J, Katayama A, Kurihara N, et al. Endocrine disruptors in the environment(IUPAC Technical Report)[J]. Pure & Applied Chemistry, 2009, 75 (5): 631–681. |
| [7] | 黄斌. 类固醇类内分泌干扰物分析方法及其在滇池水系环境化学行为研究[D]. 昆明: 昆明理工大学, 2011. |
| [8] | 胡碧波, 阳春, 张智, 等. 嘉陵江典型城市江段的类固醇雌激素分布特性[J]. 中国给水排水, 2011, 27(21): 54–58. |
| [9] | Vulliet E, Crenolivé C. Screening of pharmaceuticals and hormones at the regional scale, in surface and groundwaters intended to human consumption[J]. Environmental Pollution, 2011, 159 (10): 2929–2934. DOI:10.1016/j.envpol.2011.04.033 |
| [10] | 李本纲, 崔司宇. 中国天然雌激素排放清单和风险评价[J]. 城市环境与城市生态, 2011(4): 24–28. |
| [11] | Òvreas L, Forney L, Daae FL, Torsvik V. Distribution of bacterioplankton in meromictic Lake Saelenvannet, as determined by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S rRNA[J]. Applied & Environmental Microbiology, 1997, 63 (9): 3367–3373. |
| [12] | Webster G, Embley TM, Prosser JI. Grassland management regimens reduce small-scale heterogeneity and species diversity of beta-proteobacterial ammonia pxidizer populations[J]. Applied & Environmental Microbiology, 2002, 68 (1): 20–30. |
| [13] | Cocolin L, Aggio D, Manzano M, et al. An application of PCR-DGGE analysis to profile the yeast populations in raw milk[J]. International Dairy Journal, 2002, 12 (5): 407–411. DOI:10.1016/S0958-6946(02)00023-7 |
| [14] | 刘建林. 雌激素化合物在土壤中的吸附行为及生物降解的研究[D]. 北京: 华北电力大学, 2012. |
| [15] | 吴乔龙. 内源甾体雌激素在土壤环境中迁移转化机制研究[D]. 合肥: 合肥工业大学, 2014. |
| [16] | 石月娥. 雌激素在畜禽粪便改良农田土壤中的迁移转化及修复[D]. 长春: 吉林大学, 2016. |
| [17] | 宋晓明, 杨悦锁, 温玉娟, 等. 雌激素在地下环境中的归宿及其生态调控[J]. 中国环境科学, 2016(09): 2828–2840. DOI:10.3969/j.issn.1000-6923.2016.09.042 |
| [18] | 张照韩. 城市水体中雌激素的去除机制及复合污染效应研究[D]. 哈尔滨: 哈尔滨工业大学, 2010. |
| [19] | 王凌云. 深圳河及沿岸污水处理厂典型雌激素微生物转化研究[D]. 北京: 清华大学, 2011. |
| [20] | D'ascenzo G, Di Corcia A, Gentili A, et al. Fate of natural estrogen conjugates in municipal sewage transport and treatment facilities[J]. Science of the Total Environment, 2003, 302 (1): 199–209. |