



芝麻属于胡麻科,胡麻属,1年生草本植物,世界最古老油料作物之一。农业生产中作为栽培种的只有1种普通栽培芝麻,遗传基础很窄,故品种选育和杂种优势利用较难突破。我国自20世纪80年代后迅速发展起来的诱变育种技术,具有空间微重力、高能粒子辐射、超真空和交变磁场等特点,该技术诱发变异率高、幅度大、多数性状的遗传稳定快,能有效创制新种质,但芝麻空间诱变育种至今仍处于探索阶段[1]。
1964年国际粮农组织(FAO)和IAEA成立联合处,标志着世界原子能农业应用的正式开始。国内外开展了油料作物诱变育种研究,成果显著。印度用热中子处理晚熟蓖麻品种育成了比原品种早熟120 d、增产50%以上的新品种Aruna。我国从20世纪60年代初用γ射线对亚麻进行诱变育种,已育成黑亚四号、黑亚六号、黑亚七号、内纤亚一号4个新品种,同时获得一批优异创新资源[2]。大豆的化学诱变育种在国外始于1957年,自此各国先后开展研究,我国自1958年开始大豆诱变育种,至今已经育成一批优质、高产、抗病性强的大豆新品种。自20世纪70年代中期,人们不仅仅注重大豆产量的提高,开始转向其他方面品质的研究。Wil COX采用EMS诱变获得了含3%-4%亚麻酸的稳定突变体。1949年美国开始用X射线进行花生辐射育种,我国始于60年代,主要是山东和广东两省[3]。
将电离辐射或化学诱变手段用于改良芝麻品种,在国外始于1950年,在80年代之后进展迅速,航天诱变、杂交与化学诱变相结合、物理诱变、离子注入等方法引入了诱变育种的途径。研究较深且取得成效的是韩国、中国、埃及、印度、日本及以色列等。
根据FAO/IAEA(联合国粮农组织/国际原子能机构)的突变品种数据库(MVD)最新统计[4],截至2017年3月,全世界60多个国家在217种植物上利用诱发突变方法育成及推广了3 246个品种,其中包含中国810个,日本481个,荷兰176个,俄罗斯216个,美国139个。地区分布上的突变品种数量为:亚洲1 963个、北美洲200个、欧洲955个、非洲68个、拉丁美洲50个,澳大利亚和太平洋10个。本文就67年来芝麻诱变育种的进程简述如下。
Kobayashi[5-6]最先开始了芝麻诱变研究,并发现一些形态学及发育学的突变体。1993-1998年间,FAO/IAEA利用快中子、γ射线、NaN3和EMS得到抗病虫、闭蒴、高产、叶形变化等的突变体。Sarwar等及李英德等[7-8]用γ射线分别得到了高产抗病和隐性核雄性不育突变体。Chowdhury等[9]利用γ和X射线诱导得到2个高油的芝麻突变体,分别是短花冠和松散型分枝的突变体。Diouf等[10]用γ射线对3个遗传背景不同的芝麻材料诱变处理,得到了多毛、闭蒴、多蒴、雄性不育、开花早等突变体。另有学者利用物理化学诱变相结合、快中子及离子注入等方法进行芝麻突变体诱变研究[11-12]。我国“十五”期间“863”计划实施以来,农作物航天育种在培育新品种、保护知识产权和产业化及航天育种的机理研究等方面获得了系列重大突破。中国是世界上唯一将航天技术用于育种的国家,且空间诱变育种的研究走在了世界前列。该技术已成为快速培育优良作物品种的重要途径之一[13]。通过航天诱变技术,中国农科院油料作物研究所已成功选育2个芝麻新品种,河北省农林科学研究所粮油作物研究所已成功选育4个芝麻新品种,河南省农科院芝麻研究中心已成功选育1个芝麻新品种,本文从不同诱变因素获得的最新进展及诱变获得的芝麻农艺性状及抗病性等性状突变对芝麻的诱变成果进行综述,以期为芝麻种质创新及基因功能组学的系统研究提供理论依据。
1 芝麻诱变育种取得的成就据FAO/IAEA的突变品种数据库(MVD)最新统计,自20世纪80年代以来,截至2017年3月,8个国家诱变的芝麻突变体数量为25个。利用辐射诱变技术培育的芝麻突变体新品种数目20世纪90年代增长最为迅速。其中自2002-2005年中国利用诱变技术已育成5个芝麻突变品种,占全世界诱变芝麻突变品种比例20%,韩国6个,埃及5个,印度和伊拉克各3个。
获得突变体对于育种研究具有重要意义,是必不可少的实验育种材料。纵观遗传学发展历程,很多重大发现及突破均是有意无意从突变体的获得开始的。获得突变体的方法除了自发突变外,诱发突变也是其获得的重要途径,化学诱变及60Co-γ射线诱变、航空诱变、离子束注入等诱变途径,因其突变频率高,受到育种工作者的关注、应用并在芝麻种质创制及品种选育等方面取得了一些进展(表 1)。植物应用最多的化学诱变剂是叠氮化钠(NaN3)和甲基磺酸乙酯(EMS),均能诱发得到高密度的等位基因点突变,获得了优质、高产、抗逆、抗病等性状的芝麻突变体[14-16]。
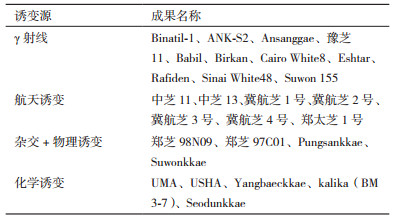
芝麻在我国分布较广泛,但容易受到环境、病虫害和芝麻本身习性的影响,因此培育出优质、高产、稳产、抗逆、抗病性强的芝麻新品种,是育种的目标。尽管传统育种技术在芝麻遗传改良方面取得了显著成就,但仍不能满足当前芝麻生产对优良品种的需求。诱变技术被广泛应用于芝麻育种中,取得了一系列重要成果(表 2)。如获得有限开花习性的突变体,节间短、分枝集中、矮杆、果粒小、花期短、抗病强、抗倒伏、花冠开裂、雄性不育等芝麻新品系[17]。
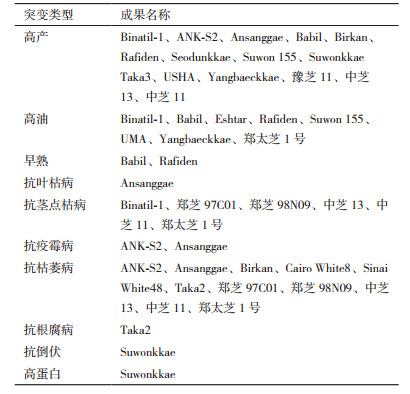
在芝麻生长过程中,花期很长且是无限花序,蒴果成熟期不一致,不适用机械收割且种子散落损失大。因此,培育节间短、矮杆、分枝集中,花期短、蒴果成熟一致的理想株型,既解决机械收割又解决了芝麻落粒的问题[18]。印度Panda等[19]在1980年经审定的名为“Kalika”的优良品种是用1% EMS处理“Binayak”栽培品种种子,在后代发现了一株矮杆、分枝紧凑的株型,这株突变体在后续多点试验中秋冬两季产量提高了15%-19%。肯尼亚的Omran[20],印度的Murty[21]也都通过诱变手段得到类似品系。高桐梅等[1]利用“实践八号”航天育种卫星搭载豫芝11、郑芝05N01、郑芝D15、日本黑芝麻、郑芝06ms030芝麻品系,对SP2和SP3代进行性状诱变研究,SP2世代,株高增加,蒴果棱数表现出六棱、八棱、牛角蒴等;茎秆出现分枝,上部有扁茎,成熟时有青色、紫色、黄色等。SP3世代中各材料后代都有数量不等分枝,不同材料在结蒴部位、蒴果长度上出现分化,蒴果棱及成熟期茎秆颜色分化同SP2世代。通过诱变的不同变异表现可选出适合通过航空诱变创造变异材料的芝麻品种。
2.2 改良种子品质运用诱发突变技术已在改良大豆、向日葵、油菜、亚麻等油料作物方面取得了成功。2004年,孟加拉国国家种子委员会注册审定的Binatil-1芝麻品种是由γ射线辐照S-30种系,随后在M2及以后世代中选择出的突变体,含油量在50%-52%之间。Danbaeckkae芝麻种子在韩国国家作物实验站于1984年经2 mmol/L的NaN3处理3 h,诱导突变得到了Yangbaeckkae品种,比对照品种Chinbaeckkae含有更高的含油量和亚油酸含量,更高的成熟率及更多的蒴果。1992年,批准的Suwonkkae芝麻品种是通过200 Gy含量的X射线照射本地的Kyum品种种子后与早期俄罗斯的抗病突变体ME-93-4杂交产生的,具有抗倒伏、高蛋白含量,抗病性好等特点。Babil和Rafiden是伊拉克本地芝麻品种分别经50 Gy和40 Gy含量的γ射线辐照产生的突变体,早熟、高油、高产。赣芝9号是我国首个通过γ射线辐射诱变选育出的黑芝麻品种,系江西省农科院作物研究所利用武宁黑芝麻60Co-γ辐射系统选育而成,具有高产、优质、抗逆性强特点[22],2008-2009两年品比试验平均产量1 520.25 kg/hm2,比对照金黄麻增产22.55%。郑太芝1号由河南省农科院芝麻研究中心用空间诱变与杂交育种相结合选育,属油用型新品种,增产潜力大、抗性优良、品质优良,抗倒性强且耐渍、耐旱,是高抗广适型芝麻品种[23]。
2.3 改良抗病性芝麻的主要病害有芝麻枯萎病、茎点枯病、疫病、青枯病等,这些病害主要靠土壤传播,在芝麻栽培种植中,主要依靠农业防治和药剂防治,改良抗病性一直是育种家的努力方向之一。Ansanggae是由早期的俄罗斯品种经200 Gy的γ射线辐照产生的突变株系,高产、抗叶枯、疫霉和枯萎病。近年来我国在芝麻诱变育种上也取得了重要进展。
中国农科院油料作物研究所利用航天诱变技术2005年选育出的中芝13具有高抗病、高耐渍、高产及广适应性的特点。在鄂豫皖赣4省生产试验中,12个试验点全部增产,平均亩产75.95 kg,比对照增产13.38%。该品种品质优,外观品质好,区试平均含油量56.58%,比对照高0.56%。2003年审定的中芝11抗倒伏能力较强,高抗芝麻茎点枯病,该品种茎点枯病发病率14.16%,病情指数5.53,比对照豫芝4号低47.12%和60.39%,抗枯萎病,耐渍,抗倒伏能力强。中芝13、中芝11抗茎点枯病及枯萎病。河南省农业科学院张海洋博士利用杂交与诱变结合的方法,经多年系谱法选育出的优质高蛋白出口型芝麻新品种郑芝98N09高抗茎点枯病,枯萎病,抗旱耐涝性强。郑芝97C01是通过杂交与辐射育种相结合,苗期粗毒素抗病性鉴定与多代连续多元病圃选择而成,产量高、稳产性好,该品种高抗茎点枯病和枯萎病,且抗旱耐渍、抗倒伏性好。河北省农林科学研究所粮油作物研究所利用航天育种培育的冀航芝2号在2010-2011年国家芝麻品种区试中抗病性表现突出,枯萎病及茎点枯病发病率均低于对照冀芝1号[24]。选育的冀航芝3号茎点枯病发病率在2011-2012年国家芝麻品种区试中发病率最低[25]。为了改良现有芝麻品种的丰产性和抗病性,以早熟、优质白芝麻品种冀芝1号纯合体为亲本,利用航天诱变育种技术,采用系统选育方法进行芝麻新品种选育,最终选育出高产、抗病芝麻新品种冀航芝4号。该品种茎点枯病和枯萎病的发病率均低于对照品种冀黑芝2号,抗病性较高[26]。1995年正式批准的斯里兰卡突变品种ANK-S2是通过200 Gy含量的γ射线辐照MI-1种子产生的,抗疫霉病、枯萎病及其他疾病。
3 近期内芝麻诱变育种目标我国是全世界芝麻基因资源最为丰富的国家之一,且我国已建立芝麻种质资源的数据库,开发及利用这些基因资源将会极大的推动我国及世界芝麻产业的发展。我国芝麻育种已走在世界的前列,但与小麦、水稻等其他作物相比仍有差距。在广泛收集、引进特异性状资源的种质资源时,还需进一步利用及评价现有保存的资源,加大搜寻光温敏雄性不育、核质互作雄性不育材料。新品种选育过程中以优质、抗逆、抗病、丰产为总的目标,继续培育强耐渍、高抗枯萎病和茎点枯病;强耐旱、耐低温、适宜秋季栽种;高含油的油用型;具特色的黄芝麻、黑芝麻保健型等特色的新品种[17]。
4 芝麻诱变育种前景展望目前,仅有非常有限的部分芝麻资源库中的材料被利用,更丰富的遗传材料还需要通过大规模种质创新来获得。芝麻种质资源的创新有远缘杂交、离子注入、物理化学诱变、航天诱变和群体改良等技术手段[27]。但芝麻品种改良诱变育种工作仍然任重道远,芝麻是二倍体作物,从庞大的M2代群体中开始选择,生育期、分枝、株高等生长特性易于简便高效率的进行观察和选择,而脂肪酸、含油量等有关品质性状则需要更大的工作量和时间才能完成检测。
国内外对理化诱变对农作物的作用机理研究不断深入,诱导变异的方向不可控,有益突变率低等问题同时存在。创制各种类型突变体,研究突变体在品质、抗逆、农艺性状等方面与未处理对照的差异,构建突变体库,与通过诱变获得新的性状类型,为研究功能基因组准备丰富的多样性材料及为遗传育种提供新的种质资源。因此,诱变技术与细胞工程、染色体工程、单倍体育种、杂交育种工程相结合,将具有更广阔的应用前景。
| [1] | 高桐梅, 卫双玲, 张海洋, 等. 芝麻空间育种后代诱变效应研究[J]. 河南农业科学, 2011, 40(4): 66–69. |
| [2] | 乔广军. 诱变在我国亚麻育种及资源创新中的利用[J]. 中国麻业, 2006, 28(1): 17–20. |
| [3] | 邱庆树. 花生辐射诱变遗传育种的研究概况[J]. 核农学通报, 1989(6): 280–283. |
| [4] | FAO/IAEA Mutant Variety Database. http://mvgs.iaea.org/. |
| [5] | Kobayashi T. Radiation genetics of sesame. PartⅢ. Morphological changes and mutants induced by ionizing radiations[J]. Japanese Journal of Genetics, 1958 (33): 239–261. |
| [6] | Kobayashi T. Radiation-induced beneficial mutants of sesame cultivated in Japan[M]. Oxford: Pergamon Press, 1965: 399-403. |
| [7] | Sarwar G, Haq MA, Chaudhry M B, et al. Evaluation of early and high yielding mutants of sesame(Sesamum indicum L.)for different genetic parameters[J]. Journal of Agricultural Research, 2007, 45 (4): 125–133. |
| [8] | 李英德, 陈清梅. 芝麻雄性不育突变体的诱变及初步遗传研究[J]. 中国油料作物学报, 1998, 20(1): 24–27. |
| [9] | Chowdhury S, Datta AK, Saha A, et al. Radiation-induced two oil rich mutants in sesame(Sesamum indicum L.)[J]. Indian Journal of Science and Technology, 2009, 2 (7): 51–52. |
| [10] | Diouf M, Boureima S, Diop T, et al. Gamma rays-induced mutant spectrum and frequency in sesame[J]. Turkish Journal of Field Crops, 2010, 15 (1): 99–105. |
| [11] | Sheeba A, Anbumalarmathi J, Babu S, et al. Mutagenic effect of gamma rays and EMS in M1 generation in sesame(Sesamum indicum L.)[J]. Research on Crops, 2005, 6 (2): 303–306. |
| [12] | 张斕, 贾瀚超, 方湉, 等. 低能C离子注入对白芝麻生物学效应影响的研究[J]. 北京师范大学学报:自然科学版, 2006, 42(1): 95–97. |
| [13] | 杨春玲, 冯小涛. 农作物太空育种进展及其发展建议[J]. 山东农业科学, 2012, 44(10): 37–39. DOI:10.3969/j.issn.1001-4942.2012.10.008 |
| [14] | Murty GSS. Induced mutants for the improvement of sesame and hybrid seed production[C]. Proceedings, 2001:99-111. http://www-naweb.iaea.org/nafa/pbg/crp/d2_3012.html |
| [15] | Pawar N, Pai S, Nimbalkar M, et al. Induction of chlorophyll mutants in Zingiber officinale Roscoe by Gamma Rays and EMS[J]. Emirates Journal of Food and Agriculture, 2010, 22 (5): 406–411. DOI:10.9755/ejfa. |
| [16] | Begum T, Dasgupta T. A comparison of the effects of physical and chemical mutagens in sesame(Sesamum indicum L.)[J]. Genetics and Molecular Biology, 2010, 33 (4): 761–766. DOI:10.1590/S1415-47572010005000090 |
| [17] | 颜廷献, 乐美旺, 饶月亮, 等. 芝麻育种技术研究进展[J]. 中国农学通报, 2010, 26(18): 146–151. |
| [18] | 石淑稳. 诱变技术在芝麻品种改良中的运用[J]. 中国油料作物学报, 1991(2): 93–96. |
| [19] | Panda BS. Mutation Breeding Newsletter[M]. Vienna: IAEA, 1981: 10-11. |
| [20] | Omran A. Oil Crops:sunflower, linseed, and sesame proceedings[J]. IDRC, 1988 : 193–200. |
| [21] | Murty GSS, Bhatia CR. Inheritance of polypetalous corolla mutation in sesame[J]. Proceedings, 1990 (2): 22–30. |
| [22] | 孙建, 涂玉琴, 乐美旺, 等. 辐射诱变育成的黑芝麻品种赣芝9号DNA变异的SRAP分析[J]. 江西农业学报, 2014, 26(10): 1–4. DOI:10.3969/j.issn.1001-8581.2014.10.001 |
| [23] | 卫双玲, 高桐梅, 张海洋, 等. 优质高产芝麻新品种郑太芝1号的选育[J]. 河南农业科学, 2016, 45(12): 49–51. |
| [24] | 徐桂真, 王生辰, 朱东旭, 等. 芝麻新品种冀航芝2号高产性稳产性及适应性分析[J]. 河北农业科学, 2013, 17(4): 59–61, 64. |
| [25] | 朱东旭, 徐婧, 武玉华, 等. 高产优质芝麻新品种冀航芝3号的选育及配套栽培技术[J]. 河北农业科学, 2015, 19(5): 54–56. |
| [26] | 徐桂真, 徐婧, 关中波, 等. 高产抗病芝麻新品种冀航芝4号的选育及适应性分析[J]. 河北农业科学, 2016, 20(6): 77–80. |
| [27] | 郑永战, 刘艳阳, 张海洋, 等. 芝麻种质资源研究进展与展望[J]. 河南农业科学, 2011, 40(6): 21–27. |