




2. 内蒙古农业大学林学院, 呼和浩特 010018;
3. 内蒙古师范大学生命科学与技术学院, 呼和浩特 010022;
4. 内蒙古和盛生态科技研究院有限公司, 呼和浩特 011517
2. Forestry College of Inner Mongolia Agriculture University, Huhhot 010018;
3. Life Science and Technology College of Inner Mongolia Normal University, Huhhot 010022;
4. Inner Mongolia and Ecological Science and Technology Research Institute Co., Ltd., Huhhot 011517
大青山自然保护区虽然具有丰富的物种资源和显著的生态功能,但是地处森林向草原的过度地带,为生态脆弱带,各物种之间及物种与环境之间的依存关系十分密切而敏感,同时,保护区植被类型略为单一,其生态系统的稳定性易遭到破坏。近些年,由于保护区植被遭到人为破坏,乱砍滥伐严重,以及保护区本身的生态脆弱性,水土流失严重。土壤作为植被赖以生存的基础,其质量直接关系到保护区生态功能的稳定[1]。而且,大青山的山地森林由于其特殊的地理构造和地形的水热再分配过程,其气候特征与所处环境中其他的生态系统有明显的不同,明确其土壤微生物丰度及土壤酶的季节动态可以更好地了解该生态系统中生物的活动规律。土壤微生物作为土壤环境中重要的生命有机体,能调控有机质和营养元素的分解和转化,是评价土壤质量的一个重要指标。不同树种处理改善了土壤微生物的生存环境,使得土壤细菌真菌群落结构明显不同。土壤酶来自于植物根系和微生物的细胞分泌物及动植物残体分解物,它能够改变土壤中矿质营养的存在状态,影响植物可利用营养的数量,在土壤物质循环中发挥着不可替代的作用[2]。微生物在其生命活动过程中,向土壤分泌大量的胞外酶。微生物死亡后,由于细胞的自溶作用把胞内酶也释放至土壤中,推动土壤生物化学反应。土壤酶是土壤中的生物活性物质及生物化学过程的积极参与者,是联系“植物-土壤酶-土壤养分”的联系纽带,在土壤生态系统的物质循环和能量转化中具有重要作用,能较客观地反映土壤质量的变化状况,间接影响到土壤中碳、氮、磷等元素的循环。庞学勇等[3]研究表明,人工林下微气候环境不同,能促进土壤养分恢复,酶活性也会增加。通过研究不同树种或植被下土壤酶活性及养分有效性的关系将有助于提高树种对土壤肥力的改善认识,同时也有助于低效人工林的结构调整和土壤肥力恢复,对管理和调控人工林和土壤生态恢复有着重要的意义。土壤酶活性在一定程度上反映了微生物的活性及其在土壤养分循环过程中的作用[4]。因此,土壤酶和土壤微生物是指示土壤理化性质微小变化的敏感指标,可以用来及时准确地反映土壤质量[5]。相较于土壤微生物量及酶的研究[6-7],对不同树种土壤微生物丰度和酶活性的关系,报道鲜少。本研究拟对大青山不同树种根围土壤微生物群落丰度和土壤酶活性与环境因子进行精确分析和科学评价,以阐明不同植被间土壤微生物群落丰度及土壤酶活性的关系,旨为改善大青山土壤生物化学环境质量寻求科学的理论依据。
1 材料与方法 1.1 材料 1.1.1 研究区概况大青山自然保护区(40°34’-40°14’ N,109°47’-112°17’ E)位于内蒙古自治区中西部包头市、呼和浩特市至乌兰察布市卓资县一线以北的阴山山地,地处内陆北纬中温带,属典型的大陆性半干旱季风气候,年平均降水量为424.6 mm,降水时间多集中在5-9月,降水量占全年总量的83.0%。年均无霜期山南平均为130-140 d,山北为90-110 d。保护区内的山区海拔为1030-2838 m。保护区随着海拔增高,植被类型依次从灌丛草原、森林灌丛草原、森林草原、向山地草原演变;地带性土壤由上而下呈带状分布,依次为山地草甸土、灰色森林土、灰褐土、栗钙土[1]。本研究选择的5个树种为白桦(Betula platyphylla)、锻树(Tilia monolica)、蒙古栋(Quercus mongolica)、虎棒子(Ostryopsis davidiana)、绣线菊(Spiraea sp.)。
1.1.2 样品采集土壤样品采自大青山小井沟森林土壤,按常规的采样方法[8],综合考虑地形地貌、植被类型、优势种、植物群落结构、人为干扰程度、恢复年限等因素,于5月中旬(春季)、9月下旬(秋季)采集不同树种(白桦、锻树、蒙古栋、虎棒子、绣线菊)的根围土样为研究对象,采用土钻法,按“S”型取土壤样品。每个树种取5颗作为重复(胸径为15 cm左右),每颗树选取3个方位,取样深度为0-30 cm的土层,将每个树种的15个样品同层均匀混合。土壤采集后除去植物根和石头,置于无菌塑封袋,新鲜土样过2 mm筛,分别于4℃、-20℃保存。
1.2 方法 1.2.1 土壤理化指标和酶活性本研究集中测定理化指标:重铬酸钾-外加热法测土壤有机碳含量;凯氏定氮法测定全氮;氯仿熏蒸浸提法测生物量碳氮含量[9]。
β-葡萄糖苷酶(βG)、N-乙酰基氨基糖苷酶(NAG)和过氧化物酶(Pod)3种酶活性,采用96微孔板法[10]。
1.2.2 土壤总DNA提取与荧光定量PCR分析土壤微生物总DNA提取使用(MOBIO Power Soil DNA Isolation Kit)试剂盒。以细菌的16s rRNA基因和真菌的ITS基因为目标基因,细菌引物为1369F/1492R,真菌引物为ITS1F/ITS4,土壤样品DNA为模板进行PCR扩增,反应体系为25 μL,其中含有12.5 μL的SYBR Green,上、下游引物(20 pmol/μL)各1 μL,模板DNA 1 μL,用灭菌的去离子水补足至25 μL。反应具体信息如表 1。
所有数据统计均在SPSS 19.0软件中进行。采用单因素方差分析法(one-way ANOVA)对数据进行方差分析和差异显著性检验(P=0.05)。采用Pearson相关系数分析指标间的相互关系。应用主成分分析法提取累计方差贡献率≥85%的公因子,所有数据均为3次重复的平均值。
2 结果 2.1 不同树种土壤理化性质的变化不同季节5个树种土壤的基本的理化性质如表 2所示。同一季节(春季或秋季)土壤有机碳、全氮、微生物量碳、微生物量氮含量和碳氮比值在不同树种间均有显著性的变化。在春季白桦树种的理化因子值均显著高于其他树种。春季和秋季绣线菊的有机碳值均为最小,分别为21.9 g/kg和35.9 g/kg。
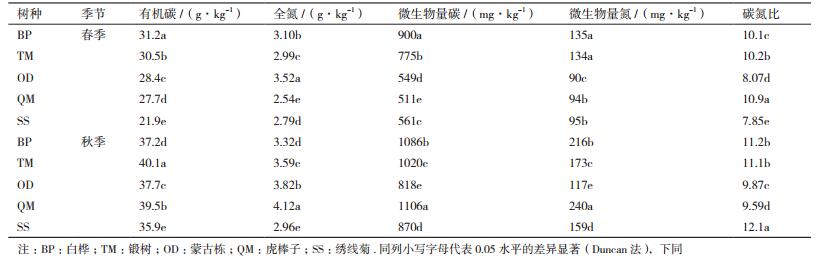
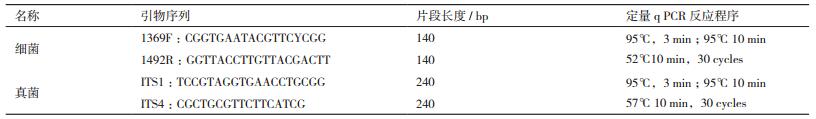
本研究的3种土壤酶活性的变化,它们在5个不同树种均有显著性的变化趋势(图 1)。在春季,β-葡萄糖苷酶这种酶活性蒙古栎最高,是183.23 nmol/(g·h),与其他树种差异显著,且在春季表现为QM > BP > OD > SS > TM。但是在秋季不同树种间差异较小,β-葡萄糖苷酶的含量在66.33-83.25 nmol/(g·h)之间。在不同季节(气温回升树木萌动期的五月与雨水充沛树木生长期的九月相较),大部分表现为春季大于秋季。而蒙古栎βG酶活性春季与秋季表现出较大的差异,春季βG酶活性是秋季的2.39倍;其余树种差异不大,在1.08-1.38倍之间。

|
| 图 1 不同树种土壤酶活性的变化 不同字母表示差异显著(LSD,P<0.05).A:β-葡萄糖苷酶活性;B:过氧化物酶活性;C:N-乙酰氨基葡萄糖苷酶 |
过氧化物酶活性在同一季节不同树种间均有显著性的变化趋势。春季和秋季的过氧化物酶活性中绣线菊都是最高的,在742-820 nmol/(g·h)之间;蒙古栎的酶活性均为最低,在456-504 nmol/(g·h)间。但是从春季到秋季,5个树种全都表现为春季的酶活大于秋季,与β-葡萄糖苷酶一样。
N-乙酰基氨基葡萄糖苷酶酶活在同一季节(春季或秋季)不同树种之间有显著性变化,在春季白桦的酶活性最高,是88.46 nmol/(g·h);在秋季蒙古栎的酶活性最高,是212.40 nmol/(g·h);在季节动态上,从春季到秋季相同树种呈现升高的趋势,尤其蒙古栎差异最大,春季NAG酶活为86.57 nmol/(g·h),而秋季之后增幅高达145%。
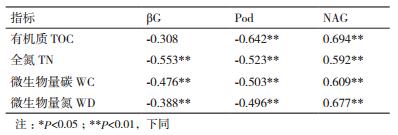
表 3所示,这两种酶不仅在树种和季节上有明显的变化,而且β-葡萄糖苷酶活性和过氧化物酶活性与全氮、土壤微生物量碳、氮含量呈极显著负相关(P < 0.01),与土壤全氮有最大的相关系数(R=-0.553)。过氧化物酶活性与有机碳也呈极显著负相关(P < 0.01),表现出最大相关系数(R=-0.642)。影响NAG酶活与土壤氮循环的的理化指标较多,均与有机碳、全碳、微生物量碳、氮呈极显著正相关(P < 0.01)。
不同季节5个树种土壤微生物群落丰度如表 4所示。不同季节,内蒙古大青山森林植物根围土壤细菌、真菌群落丰度均为秋季高于春季。其中,土壤中细菌16s rRNA基因数量最高可达14.1×106copies /mL,真菌ITS基因数量最高可达8.91×106copies /mL。
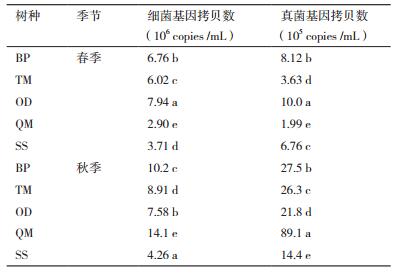
同一季节(春季或秋季)不同树种可显著改变土壤微生物群落丰度。在春季细菌、真菌群落丰度均有显著性的变化。其中细菌16S rRNA基因数量OD > BP > TM > SS > QM,真菌ITS基因数量OD > BP > SS > TM > QM;在秋季细菌16S rRNA基因数量和真菌ITS基因数量均为QM > BP > TM > OD > SS。
对于同一树种土壤微生物群落丰度均有显著的变化趋势。其中蒙古栎根围土壤细菌、真菌群落丰度在春季与秋季的差异最大,秋季细菌16S rRNA基因拷贝数是春季的4.78倍;秋季真菌ITS基因拷贝数是春季的45.42倍。绣线菊根围土壤中细菌、真菌群落丰度在春季与秋季表现出较小的差异,分别为0.87、2.16倍。但虎榛子细菌基因拷贝数春季大于秋季,差异相对较小。
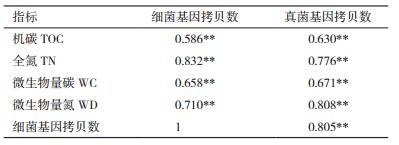
这两种微生物(细菌、真菌)群落丰度不仅在树种和季节上有明显的变化(表 5),而且与有机碳、全氮、土壤微生物量碳、氮含量呈极显著正相关(r在0-0.001之间,P < 0.01),并且细菌群落丰度和真菌群落丰度极显著正相关。
为进一步探讨影响该地区不同树种根围土壤因素变化的主控因素,对土壤酶活性、微生物群落丰度和理化因子进行主成分分析(图 2),以便能筛选出产生影响的主要因子群。前3个主成分的累计方差贡献率为87.99%(大于85%)。根据主成分分析原理,当累积方差贡献率大于85%时,即可用于近似反映系统全部的变异信息。因此前3个主成分能完全反映不同树种土壤微生物群落系统的变异信息。3个主成分中第一主成分的方差贡献率达到58.79%。第二主成分贡献率为17.99%,主要是由β-葡萄糖苷酶土壤酶活性和碳氮比决定的。第三主成分贡献率为11.20%,由过氧化物酶酶活性决定。通过上述主成分分析表明,土壤酶活性、土壤微生物群落丰度和土壤理化性质三者可以反映不同树种土壤养分的组成情况,其中有机碳、全氮、微生物量碳、微生物量氮、细菌群落丰度、真菌群落丰度、N-乙酰氨基葡萄糖苷酶等在各自的主成分中都是主导因素,可作为影响大青山不同树种根围土壤养分组成的重要因子。

|
| 图 2 基于土壤活化因子的主成分分析图 SOC:有机碳;TN:全氮;WC:微生物量碳;WD:微生物量氮;BAC:细菌群落丰度;FUN:真菌群落丰度;NAG:N-乙酰氨基葡萄糖苷酶;βD:β-葡萄糖苷酶;Pod:过氧化物酶;R:碳氮比 |
本次研究不同季节5个树种间理化因子差异性显著,可能其根系分泌物和枯落物的质和量就不同,会使得土壤物理化学性质有差异,此外,森林生态系统的季节气候变化,土壤温度、湿度的季节性变化是调控土壤微生物生物量季节波动的主要因子,致使土壤中C、N循环受到影响,而表现出上述的差异和结果。土壤氮含量的增加有助于促进植物生长,提高植物凋落物的数量和质量。而植物凋落物是土壤有机碳输入的主要来源。因此,白桦或蒙古栎有助于增加土壤有机碳的含量,促进碳的库存[11]。林下植被清除所引起的光照、土壤温度和湿度等林内条件的改变[12-13],在一定程度上间接地影响了土壤碳储量。值得注意的是,土壤碳氮比(C/N)通常被认为是土壤质量的敏感指标,可衡量土壤C、N营养平衡状况,低的C/N比可以加快微生物的分解和氮的矿化速率,在森林土壤中,高的碳氮比则使真菌比例相对较高,体现了真菌的竞争优势[14]。本次实验这5个树种土壤碳氮比均小于15,说明可使土壤中的微生物加速分解。不同季节,5种树种土壤有机碳、全氮、微生物量碳、微生物量氮均表现为秋季高于春季,这与漆良华等[15]研究的毛竹林土壤微生物量碳动态结论一致。表明不同植被类型与环境因子共同作用使得土壤碳氮含量产生差异,成为影响不同植被土壤微生物生物量差异的因素之一。随着森林植物生长和土壤环境条件的改变,土壤中可利用碳和养分资源对土壤微生物生长限制程度也在改变,进而影响土壤微生物生物量的季节变化。土壤有机质作为一种稳定而长效的碳源物质,对土壤肥力保持起着重要作用,其含量在很大程度上影响着土壤结构的形成和稳定性、植物营养的生物有效性以及土壤的缓冲性能和土壤生物多样性[16]。前人的研究报道表明,森林管理措施,如抚育、间伐等对土壤碳循环及碳的固存影响较大[17-18]。
3.2 树种对土壤酶活性的影响本实验研究了3种土壤酶活性随季节的变化,它们分别与土壤碳循环和氮循环密切相关。5个不同树种的土壤酶活性均有显著性的变化趋势。β-葡萄糖苷酶和过氧化物酶是与碳循环密切相关的土壤酶。β-葡萄糖苷酶是一种重要的水解酶,它们能够水解1-4糖苷键,释放出β-葡萄糖[14];并且裂解的二聚糖和多聚糖及β-葡萄糖苷中的β-葡萄糖苷键[19]也参与C循环[9]。过氧化物酶能氧化土壤中的有机质,能反映土壤腐殖质的强度大小。N-乙酰基氨基葡萄糖苷酶是与氮循环密切相关的土壤酶。N-乙酰基氨基葡萄糖苷酶参与氮循环中几丁质和肽聚糖的降解[20]。在森林土壤微生物环境中,土壤酶活性含量的差异主要受土壤生态条件、植被恢复阶段、土壤微生物等多因素影响。本研究秋季蒙古栎NAG酶的活性远远高于春季,但是白桦的NAG酶活性则相反,导致不同树种根围土壤这种差异的原因可能与土壤具体的理化性质以及植物分泌物对微生物的影响有关。
本研究的土壤理化因子与酶活性有很显著相关性,说明不同树种间季节相对于土壤物理化学性质或者树种对于碳循环相关土壤酶活性有更强的影响。土壤全氮、微生物量碳、氮含量越高,土壤中与碳循环相关的土壤酶活性越低。因为这两种酶从春季到秋季树种的酶活性呈下降趋势,可能由于气候、水分等条件的影响。土壤酶的时间动态变化主要由于季节性的环境因子变化导致的,并且与植物生长周期变化密切相关。季节的变化引起土壤生态系统中水热等条件的改变,这些变化影响土壤酶的分解、土壤酶的周转以及植物对土壤酶的利用。土壤中各种酶之间存在着相互促进而又相互竞争的关系,各种土壤酶的不同步分解是温带森林生态系统中养分保留和生产力保留的一种重要机制。不同季节,酶作用方式不同,群落结构和功能也存在差异,从而表现出了酶的季节动态格局[21]。土壤物理化学性质相对于季节或不同树种对与氮循环相关土壤酶活性有更强的影响。土壤有机碳、全氮、微生物量碳、氮含量越高,土壤中与氮循环相关的土壤酶活性越高,这与斯贵才等[14]研究发现藏东南森林土壤微生物群落结构与土壤酶活性的结果一致。
3.3 树种对土壤微生物群落丰度的影响土壤微生物类群之间含量上的变化是土壤微生物群落结构演替的重要内容,尤其是土壤细菌和真菌生物量的变化[14],这对土壤微生物丰度有很大的影响。本实验用荧光定量PCR技术研究了大青山不同树种土壤微生物丰度的变化规律。内蒙古大青山森林植物根围土壤细菌、真菌群落丰度均为秋季高于春季。这可能是季节变化的过程中,土壤中腐殖质的含量得到相应的增加,土壤中微生物的生长得到促进的原因。其中,土壤中细菌16S rRNA基因数量最高可达14.1×106copies /mL,高于韩文炎[7]等研究的茶园土壤中16S rRNA基因数量,低于王轶等[14]研究的大田土壤中细菌16S rRNA基因数量。真菌ITS基因数量最高可达8.91×106copies /mL,高于王轶等[14]研究的大田土壤中真菌ITS基因数量,导致这种差异的原因可能与土壤具体的理化性质以及植物分泌物对微生物的影响有关。同一季节(春季或秋季)不同树种可显著改变土壤微生物群落丰度。在春季细菌、真菌群落丰度均有显著性的变化。说明树种可显著影响土壤微生物群落丰度,尤其在大青山森林植被中,相较于其他树种种植白桦可显著提高微生物群落丰度。也说明不同的树种随着季节不同,气候水热因子的改变,呈现不同的变化趋势。这也与树种的特性,根系的分泌物等有很大的关系。
这两种微生物(细菌、真菌)群落丰度与土壤理化因子呈极显著正相关,说明整个土壤微生物群落环境C、N循环具有非常重要的作用。细菌群落丰度和真菌群落丰度极显著正相关。真菌和细菌是土壤微生物的主要组成部分,真菌对枯枝落叶的分解能力极强,是进行腐解作用的主要微生物,并在土壤的生物化学转化过程中起相当重要的作用。土壤中的细菌几乎参与了土壤中的所有生物化学过程,并具有快速生长的能力,能旺盛地分解各种自然物质,故在土壤的物质转化过程中具有重要的作用。因而土壤中的真菌在参与枯枝落叶的分解过程中能有效促进土壤细菌的生长,使二者呈极显著正相关关系(r为0,P < 0.01)。该研究结果与李强等[4]的研究一致。因此,森林生态系统中不同树种是影响土壤微生物群落丰度(细菌、真菌)的重要因素。
3.4 不同树种土壤理化指标、酶活性及微生物丰度的季节变化动态关系季节变化动态对不同树种根围土壤理化指标、酶活性及土壤微生物丰度均有显著影响。季节更替能显著影响土壤温度与湿度,进而对参与碳氮循环的微生物、植被组成等产生显著影响。本研究得出了不同树种秋季的土壤理化指标、微生物群落丰度均高于春季的结论。这与李胜蓝等[22]研究的森林类型土壤微生物生物量碳氮含量中“秋高春低”型结论一致。从春季到秋季,由于气温的升高,雨水的增加更利于植物凋落物的分解,也利于微生物的生长。而酶活性在季节动态上分布有差异。大量研究表明,不同试验区酶活性最高值出现的季节不同[23],而真菌生物量表现为在水热条件相对缓和的秋季较佳,这也进一步说明三者之间的协同响应关系。
4 结论本文研究了大青山不同树种土壤微生物丰度和土壤酶活性的变化,研究结果表明,影响大青山不同树种根围土壤的因素不均一,主要表现为气候中的水热因子和树种的特性。并且,土壤理化因子与土壤微生物丰度呈极显著正相关关系,由此可推断,树种可能是通过影响土壤理化特性间接影响土壤微生物群落丰度。
| [1] | 齐云龙. 大青山古路板不同林型土壤AM真菌多样性及其分布特征的研究[D]. 呼和浩特: 内蒙古大学, 2012. http://cdmd.cnki.com.cn/Article/CDMD-10126-1012440259.htm |
| [2] | 杨成德, 龙瑞军, 薛莉, 等. 东祁连山高寒草本草地土壤微生物量及酶的季节动态[J]. 中国草地学报, 2014, 36(2): 78–84. |
| [3] | Pang X, Bao W. Effect of substituting plantation species for native shrubs on the water-holding characteristics of the forest floor on the Eastern Tibetan Plateau[J]. Journal of Resources & Ecology, 2011, 2 (3): 217–224. |
| [4] | 李强, 靳振江, 李忠义, 等. 岩溶地貌部位对土壤微生物丰度与酶活性的影响[J]. 水土保持通报, 2014, 34(3): 50–54. |
| [5] | Baddam R, Reddy GB, Raczkowski C, et al. Activity of soil enzymes in constructed wetlands treated with swine wastewater[J]. Ecological Engineering, 2016, 91 : 24–30. DOI:10.1016/j.ecoleng.2016.01.021 |
| [6] | 李世卿, 王先之, 郭正刚, 等. 短期放牧对青藏高原东北边缘高寒草甸土壤及微生物碳氮含量的影响[J]. 中国草地学报, 2013, 35(1): 55–60. |
| [7] | 牛得草, 江世高, 秦燕, 等. 围封与放牧对土壤微生物和酶活性的影响[J]. 草业科学, 2013, 30(4): 528–534. |
| [8] | 覃勇荣, 苏盛, 黄小梅, 等. 不同植被对石漠化地区土壤微生物数量的影响[J]. 河池学院学报, 2016, 36(5): 1–8. |
| [9] | 何建州, 杨金燕, 田丽燕, 等. 用紫外-荧光微孔板酶检测技术测定两种土壤的酶活性[J]. 四川农业大学学报, 2012, 30(2): 181–185. |
| [10] | 陈丽霞. 内蒙古大青山森林土壤微生物量碳氮及微生物特征研究[D]. 呼和浩特: 内蒙古农业大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10129-1015427811.htm |
| [11] | 杨丹, 余旋, 刘旭, 等. 栽培模式对沙棘人工林土壤微生物群落结构和参与氮循环功能基因的影响[J]. 应用生态学报, 2015, 26(12): 3634–3640. |
| [12] | 马芳芳, 贾翔, 赵卫, 等. 间伐强度对辽东落叶松人工林土壤理化性质的影响[J]. 生态学杂志, 2017, 36(4): 971–977. |
| [13] | Chen XL, Wang D, Chen X, et al. Soil microbial functional diversity and biomass as affected by different thinning intensities in a Chinese fir plantation[J]. Applied Soil Ecology, 2015, 92 : 35–44. DOI:10.1016/j.apsoil.2015.01.018 |
| [14] | 斯贵才, 袁艳丽, 王建, 等. 藏东南森林土壤微生物群落结构与土壤酶活性随海拔梯度的变化[J]. 微生物学通报, 2014, 41(10): 2001–2011. |
| [15] | 漆良华, 艾文胜, 范少辉, 等. 湘中丘陵区不同经营类型毛竹林土壤微生物量碳动态[J]. 南京林业大学学报:自然科学版, 2013, 37(5): 45–48. |
| [16] | 刘苑秋, 王芳, 柯国庆, 等. 江西瑞昌石灰岩山区退耕还林对土壤有机碳的影响[J]. 应用生态学报, 2011, 22(4): 885–890. |
| [17] | 吴亚丛, 李正才, 程彩芳, 等. 林下植被抚育对樟树人工林土壤活性有机碳库的影响[J]. 应用生态学报, 2013, 24(12): 3341–3346. |
| [18] | 刘迎春, 王秋凤, 于贵瑞, 等. 黄土丘陵区两种主要退耕还林树种生态系统碳储量和固碳潜力[J]. 生态学报, 2011, 31(15): 4277–4286. |
| [19] | 刘强, 彭少麟. 植物凋落物生态学[M]. 北京: 科学出版社, 2010. |
| [20] | 赵杏, 钟一铭, 杨京平, 等. 不同植茶年限土壤碳氮养分及胞外酶对干旱胁迫的响应[J]. 生态学报, 2017, 37(2): 387–394. |
| [21] | 黄黎英, 曹建华, 周莉, 等. 不同地质背景下土壤溶解有机碳含量的季节动态及其影响因子[J]. 生态环境学报, 2007, 16(4): 1282–1288. |
| [22] | 李胜蓝, 方晰, 项文化, 等. 湘中丘陵区4种森林类型土壤微生物生物量碳氮含量[J]. 林业科学, 2014, 50(5): 8–16. |
| [23] | 牛小云, 孙晓梅, 陈东升, 等. 辽东山区不同林龄日本落叶松人工林土壤微生物、养分及酶活性[J]. 应用生态学报, 2015, 26(9): 2663–2672. |