




2.江苏省兴化市农业委员会,兴化225700
2.Agricultural Commission of Xinghua City in Jiangsu Province,Xinghua 225700
在20世纪80年代我国杂交粳稻出现了迅速发展,但其种植面积所占粳稻总面积的比例仅3%左右,远小于杂交籼稻所占的70%[1]。近年来,虽然出于国家粮食安全的需要,国家及科研单位加大力度研究和推广,但杂交粳稻的发展速度仍然缓慢,究其原因杂交粳稻的低制种产量严重影响了其产业化的发展[2]。
粳稻不育系的异交结实率低是制约杂交粳稻制种产量的主要因素。田大成等[3]研究认为,柱头外露颖花与非外露颖花异交能力存在巨大差异,前者结实率比后者一般高3-4倍,依靠柱头外露颖花开花当天及以后接受花粉受精结实是异交结实的重要途径,开花当天异交结实一般不到总粒数的50%,而开花第2天后一般可占50%。李庆荣[4]认为柱头外露颖花的结实率占制种异交结实的 70%-80%,是构成制种产量的主体。杨保汉等[5]研究认为,在自然条件下,不育系柱头外露率每提高1%,结实率可提高0.74%-0.92%,每公顷可增产47-68 kg。林建荣等[6]研究表明保持系柱头外露与不育系柱头外露存在着极著的遗传关系。由浙江省嘉兴市农业科学研究院选育的晚粳稻嘉58,其对应的BT型不育系嘉58A,不育性稳定、不育度高,综合农艺性状优良,具有良好的配合力,但柱头外露低(约为1.0%),利用嘉58A配制的杂交粳稻(组合)皆因制种产量低而影响在生产上的应用。本研究试图将控制水稻柱头外露率的QTL导入嘉58A,探索利用分子标记辅助选择进行人工创制高柱头外露率粳稻不育系的技术可行性。
1 材料与方法 1.1 材料以天津市农科院提供的高柱头外露率育种材料T60B(来自50S,籼型,柱头总外露率73%)为父本,浙江省嘉兴市农业科学研究院(所)三系粳稻保持系嘉58B(柱头总外露约为1.0%)为母本进行杂交和回交,嘉隆17B和嘉隆43B是嘉58B高柱头外露率基因导入系。试验在浙江省嘉兴市农业科学研究院(所)试验农场进行,群体及系圃均采用单本植,插植规格为23.0 cm×16.7 cm,水肥管理及病虫害防治按一般生产用田。
1.2 方法 1.2.1 考察柱头外露率2014年6月,种植嘉隆17A、B系,嘉隆43A、B系和嘉58A、B系鉴定圃(各1 000株),待不育系穗杨花期随机选取100个单株的主茎穗,通过目测调查每穗的总颖花数、单边柱头外露颖花数和双边柱头外露颖花数,计算单边柱头外露率和双边柱头外露率及总外露率。
1.2.2 分子标记检测50S携带的柱头外露的QTLs连锁标记为SSR标记。检测目标QTL的水稻DNA采用赵国珍等[7]提法提取,PCR反应体系为:DNA模板2 μL,灭菌水20 μL和混合物16 μL。其中混合物为:灭菌水8.0 μL,dNTP 2.8 μL,Buffer 4.0 μL,左右引物各0.5 μL,Taq酶0.2 μL。PCR扩增程序为:94℃ 4 min;94℃ 30 s,55℃ 30 s,72℃ 40 s,35个循环;72℃ 4 min;25℃ 1 min。扩增产物用4%的琼脂糖凝胶电泳检测。
1.2.3 考察异交结实率2014年6月至10月在浙江省嘉兴市农业科学研究院(所)农场,用花时中等偏早的籼粳交恢复系HR511为父本,嘉隆43A、嘉隆17A和嘉58A为母本,进行大田小区制种。根据不育系的始播历期合理安排播种期。试验中父本HR511安排在6月13日播种,母本安排在6月6日播种,使父母本均能在9月3日左右始穗。父母本行比为2∶4,行距23.0 cm,株距16.7 cm,单本植,每个不育系制种小区面积约为30 m2,2次重复。父母本杨花后,每个小区从不育系中间2行选取整齐一致的穗子20个进行挂牌,选其中10个穗,午后观察柱头外露情况,并对柱头单双外露的颖花用油笔做好记号,每天分2个时段(10:30-11:00,11:30-12:00)进行人工辅助赶粉,直到父本的颖花开完为止齐穗后25 d,同时选取非标记穗10个,调查籽粒的柱头外露粒的柱头夹持及结实情况。采用DPS软件对调查结果进行统计分析。
1.2.4 考察外露柱头的结实率采用延迟授粉法,使父本开花时隔离,标记每天开的母本颖花(100粒),去除闭颖粒和未开花粒,然后在同一天使标记的颖花接受父本花粉,闭颖后开花标记粒的柱头外露情况,用柱头外露粒结实率作为衡量柱头活性的大小。分别于开花后0、1、2、3、4、5、6和7 d 授粉计算柱头外露粒结实率,计算公式为:某天外露柱头结实率(%)=柱头外露粒结实粒数/柱头外露粒数×100%。
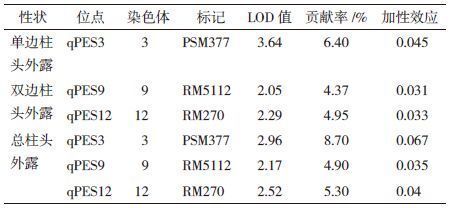
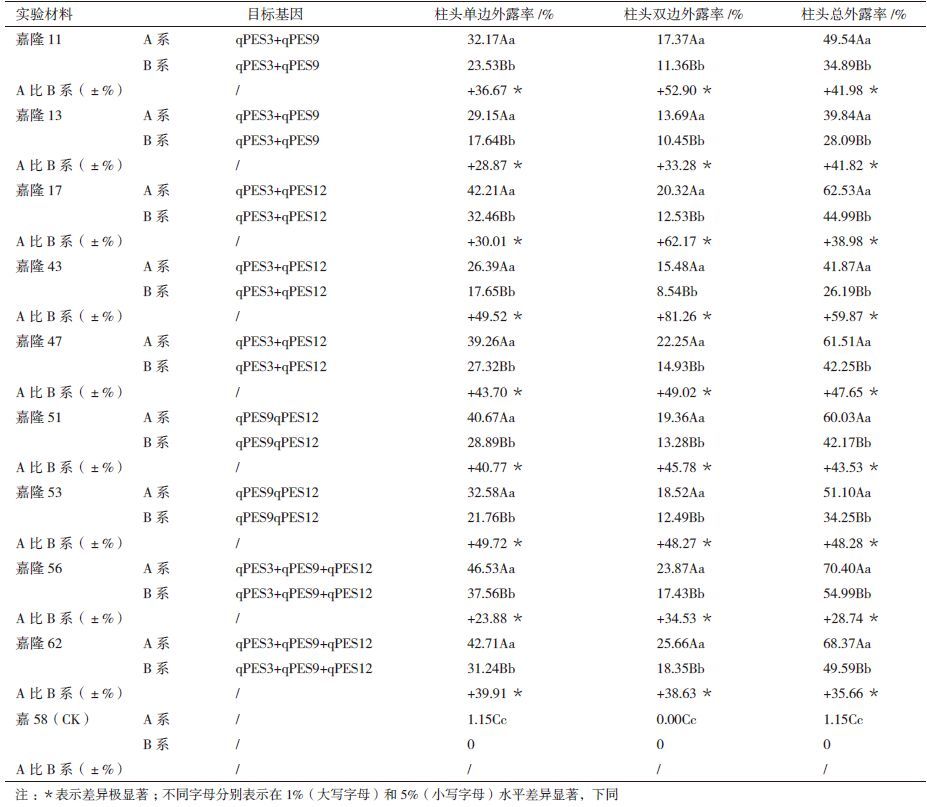
2 结果 2.1 柱头外露QTL分子检测及表型利用分子检测能够将调控柱头外露的QTL(图 1和表 1)导入粳稻品系中(表 2,部分导入系)。本试验中qPES3、qPES12连锁标记PSM377、RM270在T60B和嘉58B多态性显著(图 2),导入系嘉隆11A、嘉隆13A导入了qPES3+qPES9,嘉隆17A、嘉隆43A和嘉隆47A入了qPES3+qPES12,嘉隆51A、嘉隆53A导入了qPES9+qPES12,嘉隆56、嘉隆62A导qPES3+qPES9+qPES12;嘉隆17B和嘉隆43B早实现综合农艺性状的稳定,株型特征亲嘉58B,其不育系为嘉隆17A和嘉隆43A,经多代分子检测显示嘉隆17B、嘉隆43B均携带qPES3和qPES12位点(图 3)。表 2中列出的9个系表型值,有5系的柱头外露率超过60.0%,2系在40.0%-50.0%,2系在30.0%-40.0%,较嘉58A均表现极显著提高,且柱头外露率(qPES3+qPES9+qPES12)>柱头外露率(qPES3 +qPES12或qPES3 +qPES9)。

|
| 图 1 几个粳稻不育系的柱头外露表现 |

|
| M :标准分子量;1,3 :T60B ;2,4 :嘉58B 图 2 PSM377 和RM270 在亲本间多态性分析 |

|
| 1-12 :分离世代材料;A :qPES3 连锁标记PSM377 电泳图谱;B :qPES12连锁标记RM270 电泳图谱 图 3 柱头外露QTL(qPES3 和qPES12)的分子标记检测 |
本研究调查了嘉隆17A、嘉隆43A和嘉58A三个不育系的杨花期外露柱头籽粒和柱头发生夹持籽粒的结实情况。结果(表 3和图 4)表明,嘉隆17A的柱头单边外露籽粒结实率为71.22%,柱头双边外露籽粒为73.37%,占总结实粒的比率分别为64.01%和28.65%,二者对总结实率的贡献率超过82.0%,嘉隆43A的柱头单双边外露籽粒结实率分别为67.53%和62.69%,占其总结实粒比例53.75%和28.72%,对总结实粒的贡献率超过81.0%。


|
| 图 4 几个粳稻不育系柱头夹持情况 |
表 3同时表明,嘉隆17A单双边柱头夹持率为99.65%和97.0%,嘉隆43A分别为99.0%和97.8%,单柱头的夹持率略高于双柱头的夹持率,嘉58A柱头不发生夹持现象,嘉隆17A单双边柱头夹持粒结实率分别达75.16%和72.94%,占总结实粒的比超过85.0%,嘉隆43A表现类似规律;研究还表明嘉隆17A和嘉隆43A的夹持粒结实率均略高于其外露粒结实粒。
2.3 花后外露柱头活性的动态变化图 5显示,3个粳稻不育系的杨花当天(花后0 d)柱头活性最强,均接近80.0%的结实率,其中柱头外露不育系嘉隆17A和嘉隆43A的柱头略高于柱头非外露不育系嘉58A,在花后1-6 d,嘉58A花后1 d后柱头活性激剧下降,花后2 d后柱头基本没有活性,与嘉58A相比,嘉隆17A和嘉隆43A的柱头在花后1.5-5 d仍有较高的授粉结实能力,柱头活力下降幅度较平缓。其中花后1-5 d嘉隆43A柱头活性高于嘉隆17A,这可能与嘉隆43A双柱头外露率高于嘉隆17A,表 3中嘉隆17A和嘉隆43A的双柱头外露籽粒结实率均高于其对应的单柱头外露籽粒的结实率,进一步验证上述结论。

|
| 图 5 几个粳稻不育系外露柱头籽粒花后活性动态 |
李晨等利用栽培稻和野生稻的BC群体在第2、5和8染色体上定位到控制柱头外露率的QTL。尹成[12]和沈圣泉等[13]在考虑到基因互作和GE互作的前提下,检查出位于6号染色体贡献率达到19.1%的主效QTL,虽然以上研究发现的水稻柱头外露的QTL贡献率都达到10%以上,但是对其应用限制,还主要是对QTL的紧密连锁的分子标记发掘不够,不能高效精确实现标记辅助选择。笔者试图利用T60B携带控制柱头外露的QTL连锁标记,将qPES3、qPES12成功导入粳稻背景中并获得qPES3+qPES12的稳定表达粳稻不育系,研究结果再次证明利用分子标记可以有效辅助人工创制高柱头外露率的粳稻不育系。本研究中的两个不育系柱头外露表现差异明显,可能与人工选择背景有关,即qPES3、qPES12表达存在基因型差异,与其保持系相比,不育系的柱头外露率显著高于对应保持系,这可能因为在粳稻背景下控制柱头外露QTL也存在不育胞质效应,与籼稻中报道一致。
籼稻不育系的柱头外露颖花在开花当天及其后的3 d内都具有较强的受精能力,有花粉供给时柱头外露率在60%以上,则结实率一般不低于60%,活性柱头率98%以上,柱头活性强的不育系能够有效利用父本的非及时花粉提高不育系结实率[14]。笔者研究了粳稻不育系的杨花后柱头活力的动态变化,结果表明,粳稻不育系的柱头活性在开花当天均表现较强,活性柱头率80%左右,不育系间差异不显著,但花后1-7 d,柱头外露不育系的柱头在花后1-5 d表现较强的接受花粉结实的能力,无柱头外露不育系的花后2 d柱头活性急剧下降、3 d后基本不具备活性,与柱头外露的籼稻不育系相比,柱头外露的粳稻不育系活性柱头率表现较低,但柱头的活性持续时间较长。本研究比较了粳稻不育系外露柱头的夹持情况,结果认为柱头外露粒95%以上柱头发生了夹持现象,粳稻不育系杨花闭颖时柱头外露粒和籽粒蜡熟黄熟期柱头夹持粒表现趋势一致,在育种实践中可通过考察籽粒的柱头夹持率来选择柱头外露性状的目的。
4 结论本研究通过分子标记与常规育种技术相结合,有效改良了粳稻不育系的柱头外露率,来自籼稻的QTLs能够在粳稻中稳定表达。研究结果证实了分子标记技术在选择水稻QTLs的可能性和有效性,同时表明要提高粳稻的异交特性仍然要通过利用籼稻的有利性状来实现。
| [1] | 荆彦辉, 张子军, 周晓玲, 等. 水稻柱头外露基因的遗传及应用思考[J]. 农业科技通讯, 2009(8):40-42. |
| [2] | 龙俐华, 舒琨. 提高杂交稻异交率的研究[J]. 湖南农业大学学报:自然科学版, 2000, 26(3):167-170. |
| [3] | 田大成, 黄三奎, 段永国, 等. 水稻不育系花时和受粉时间与异交结实率的关系[J]. 杂交水稻, 2004, 19(3):50-54. |
| [4] | 李庆荣. 柱头外露率的环境变异及其在异交结实中的作用[J]. 种子科技, 1995(2):32-33. |
| [5] | 杨保汉. 不育系柱头外露率及其结实率研究[J]. 杂交水稻, 1997, 12(1):13-15. |
| [6] | 林建荣, 吴明国, 宋昕蔚. 三系粳稻不育系开花习性与异交结实率的关系[J]. 杂交水稻, 2006, 21(5):69-72. |
| [7] | 赵国珍, 贾育林, 严宗卜. 一种高效便捷的水稻DNA提取法及其应用[J]. 中国水稻科学, 2012, 26(4):495-499. |
| [8] | 冯玲玲, 荆彦辉, 黄成, 等. 水稻柱头外露率的QTL分析[J]. 北方水稻, 2010, 40(3):20-22. |
| [9] | 李晨, 孙传清, 穆平, 等. 栽培稻与普通野生稻两个重要分类性状花药长度和柱头外露率的QTL分析[J]. 遗传学报, 2001, 28(8):746-751. |
| [10] | Miyata M, Yamamoto T, Komori T, Nitta N. Marker-assisted selection and evaluation of the QTL for stigma exsertion under japonica rice genetic background[J]. Theor Appl Genet, 2007, 114(3):539-548. |
| [11] | 邓应德, 应杰政, 石媛媛, 等. 控制水稻柱头外露率的数量性状基因座初步分析[J]. 湖南农业大学学报, 2010, 36(4):373- 376. |
| [12] | 尹成, 李平波, 高冠军, 等. 水稻柱头外露率QTL定位[J]. 分子植物育种, 2014, 12(1):43-49. |
| [13] | 沈圣泉, 庄杰云, 包劲松, 等. 水稻柱头外露率QTLs定位及其互作分析[J]. 生物数学学报, 2006, 21(4):610-614. |
| [14] | 李楠. 水稻不育系外露柱头存活率测定及提高措施[J]. 湖北农业科学, 2013, 52(3):514-517. |