




2. 山东省泰安市林业局,泰安 271000
2. Tai’an Forestry Administration,Tai’an 271000
园林废弃物是指园林绿化植物自然凋落或人工修剪、园艺操作过程中所产生的枯枝、落叶、草屑、花败、树木与灌木剪枝以及其他植物残体[1]。随着城市化进程的加快以及城市园林绿地面积的持续增加,园林废弃物成为城市固体废弃物的重要组成之一,给环境保护和可持续发展带来巨大的压力。园林废弃物中包含大量木质纤维成分,目前处理的方法主要有直接填埋、焚烧、粉碎粗回收、热化学处理、生产垃圾燃料及直接堆放腐熟等,常常造成二次环境污染和巨大的资源浪费。如何合理高效地将其转化利用已成为当前研究和资源开发应用的热点和难点。
利用微生物高效降解转化的堆肥化技术具有成本低、无污染、产品肥效长和利于土壤改良等优点,能够充分实现废弃物的无害化、减量化、资源化利用[2]。通过施加高效降解的微生物菌剂对园林废弃物进行生物好氧发酵处理,可以大大缩短腐熟时间、增加肥效、减少环境污染和安全隐患,实现资源的高效再利用,对环境的可持续发展和资源的循环利用具有重要意义。
1 微生物堆肥技术中的功能微生物区系高效降解的微生物菌剂在农业堆肥生产中已得到广泛应用,但在园林废弃物堆肥中的功能和作用尚有待于进一步研究和开发。园林废弃物、农业废弃物以及部分城市生活垃圾主要是由纤维素、木质素、半纤维素、蛋白质、脂类等按不同比例组成的混合物,含丰富的有机物和植物生长所需的营养元素如N、P和K及微量元素Cu、Zn、Fe和Mn等。微生物堆肥处理过程实质上是微生物将复杂的有机物分解转化成植物可以吸收和利用的营养成分。根据反应过程不同阶段对氧气需求的不同,常将堆肥分为好氧和厌氧两种。厌氧堆肥主要分为产酸和产甲烷两个阶段,产生的氨气、硫化氢以及其他还原性气体易挥发产生恶臭气味具有较高热值,但需要专用发酵设备以便收集,设备投资较大。利用微生物进行好氧发酵为研究重要方向,按照分类和功能的不同,好氧发酵中的微生物主要可分为如下几类。
在好氧发酵系统中,细菌是重要微生物区系,因其菌体较小,具有较大的比表面积,可迅速将可溶性底物吸收到细胞中以生长繁殖,数量明显多于真菌。不同堆肥环境中分离的细菌也存在差异,例如,假单胞菌属(Pseudomonas)、克雷伯氏菌属(Klebsiella)和芽孢杆菌属(Bacillus)等是重要的解脂肪菌,而芽孢杆菌属同时还具有高效降解蛋白质、淀粉的功能,是有机物的重要分解者。堆肥发酵腐熟是一个逐渐升温的过程。发酵初期,嗜温细菌作为主要的优势菌群,随着温度升高,嗜热细菌则随之增多,成为堆肥高温阶段的优势菌群[3]。其中多数为芽孢菌,如枯草芽孢杆菌(B. subtilis)和地衣芽孢杆菌(B. licheniformis)等,该类细菌因能形成很厚的芽孢壁而耐受高温、腐蚀及营养物缺乏等不良环境条件,其在80℃左右的堆温下生长繁殖,是高温堆肥腐熟降解的主要菌群。
放线菌主要利用半纤维素并能溶解木质素类复杂有机物,在高温下分解纤维素、角质素等。放线菌在园林废弃物堆肥技术中可用于树皮等坚硬物质的分解,作用甚广。链霉菌(Streptomyces)、诺卡氏菌(Nocardia)及高温放线菌(Thermoactinomyces)等都是堆肥高温阶段中常见的嗜热放线菌,在堆肥过程高温阶段对木质纤维素以及其他物质的分解起着重要作用[4]。
真菌中的白腐菌、褐腐菌对堆肥中极难降解的木质纤维具有高效降解作用。真菌可通过其分泌的胞外酶,促进有机物的生物降解。如木霉(Trichoderma)能降解纤维素,曲霉(Aspergillus)能够降解果胶、纤维素等,假丝酵母菌(Candida)能降解半纤维素等,但在高温堆肥环境中可以耐受一定高温的真菌才能高效降解木质纤维素。因此,高温真菌对于堆肥物料中的木质素、纤维素等的分解转化具有十分重要意义。
另外,堆肥中的功能微生物还包括固氮、解磷、解钾、生防及促生菌等,它们能有效促进堆肥中碳、氮循环,将土壤中难以被植物吸收的矿物质转化为植物可吸收的营养元素,增加土壤肥力,还林还田后对植物的生长、抗逆性、生物防治等都具有明显作用。因此,通过了解堆肥成分及好氧发酵过程的动态变化,筛选获得合理的功能微生物进行处理,促成有机物的有效动态分解,将其转化为稳定的腐殖质是实现农林废弃物资源化利用的重要途径[5]。
2 园林废弃物堆肥化过程中微生物的功能与作用微生物是堆肥过程中最重要的生物因素,采取适合的调节措施促进堆肥微生物的生长代谢可能提高堆肥效率与产品质量。在微生物菌剂中,主要微生物菌种的功能及应用领域统计,如表 1所示。根据园林废弃物的组成特点以及功能微生物在堆肥中的作用,对其进行微生物堆肥化处理,应着重考虑以下几个方面。
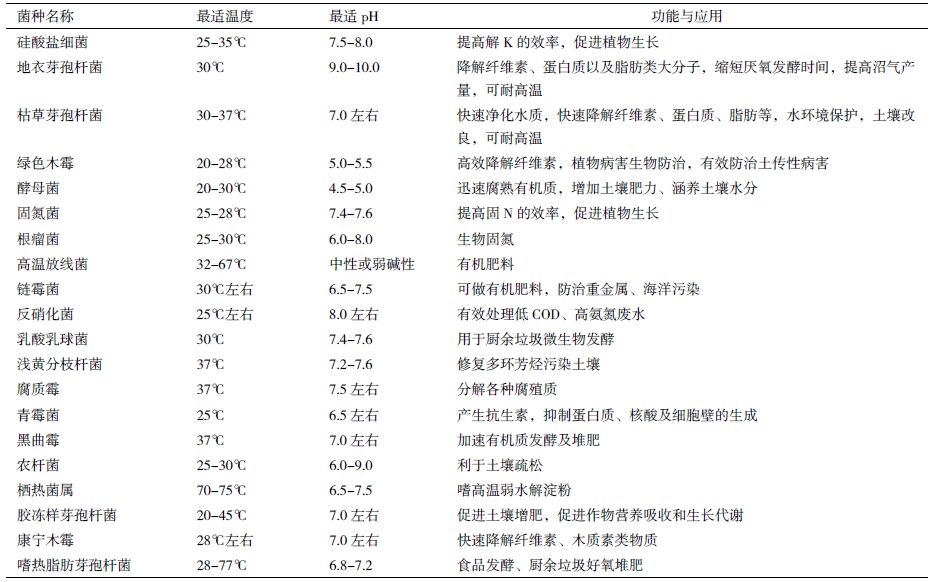
园林废弃物含有大量的有机木质纤维碳物质及一些可溶性糖、有机酸和淀粉等。其中,木质纤维主要包括木质素、纤维素和半纤维素,在自然条件下降解非常缓慢,是制约园林绿化废弃物高效转化的重要因素。降解园林废弃物中的木质素、纤维素,缩短堆肥腐熟时间、提高堆肥的生产效率,使其成为循环再利用的物质是接种微生物菌剂的首要目的[6]。
植物枯枝、落叶等天然木质素的完全降解通常依赖于自然界真菌、细菌、放线菌及其产生的多种降解酶系的共同作用,其中真菌起着主导作用[7]。白腐菌是目前公认的最高效的木质素降解微生物,不产生色素,能在木质材料中形成白色腐朽残余物,降解木质素的能力远高于其降解纤维素的能力[8]。白腐菌与褐腐菌、软腐菌等共同作用,其中,褐腐菌和软腐菌将木质纤维基质中的纤维素分解,白腐菌再通过分泌相应的木质素降解酶将大量木质素降解。研究最多的木质素降解酶有3种,即木质素过氧化物酶(LiP)和锰过氧化物酶(MnP)及漆酶(Laccase)[9]。LiP催化通过自由基调节C-C键的断裂造成木质素分子中Ca-Q3键断裂,从而分解木质素结构;MnP广泛存在于白腐菌中,主要依靠催化Mn2+ 转为高度活性的Mn3+,利用Mn3+去氧化木质素酚型结构,使α碳原子被氧化、烷基苯基键断裂、C-C键断裂木质素裂解;Lac在酚类底物存在的情况下,能由Mn2+产生Mn3+螯合物,并在Lac和MnP两种酶协同作用下有效降解木质素[10]。
在纤维素降解微生物中,真菌种类也明显多于细菌和放线菌,如木霉属能分泌丰富的纤维素酶系并具有极高的酶活力[11]。纤维素酶是多酶组分的一个复杂酶系,主要分为外切葡聚糖酶(CBH)、内切葡聚糖酶(EG)和β-葡萄糖苷酶(CB)3大类[12]。协同理论目前被大多数学者普遍接受,该理论认为:纤维素降解是由多种酶共同作用的结果[13]。内切葡聚糖酶首先进攻纤维素的非晶区,形成游离末端,然后外切纤维素酶从多糖链的非还原端切下纤维二糖单位,β-葡萄糖苷酶再水解纤维二糖单位形成葡萄糖[14]。但协同理论并没有对该作用的起始反应机制进行说明,特别是对结晶区的降解机制没有很好的解释。
木质纤维是自然界最大的可再生的生物质资源和能源,也是园林废弃物的主要组成部分。因此,了解木质纤维的降解机理筛选高效的降解微生物是园林废弃物堆肥化技术中的关键因素之一。
2.2 参与氮素循环微生物氮素是土壤肥力和植物生长的主要成分。园林废弃物虽然含有丰富的有机物及碳元素,但其中的氮素成分含量却很少,无法为植物提供大量的氮素营养。堆肥化过程伴随着有机氮的矿化(NH3 的挥发)、硝化及反硝化作用(释放N2O),同时也进行着无机氮的生物固定作用,各种作用相互转化相互依存,都是复杂的微生物活动过程。这个过程最大的负面影响是氨气挥发,导致氮素损失、降低堆肥肥效[15]。氮含量的减少同时也意味着营养元素的损失,研究堆肥中的氮素微生物生理群的变化有助于了解氮素转化、控制氮素形态和损失情况[16]。
堆肥中氮的形态主要有铵态氮、硝态氮和有机氮,其中有机氮占总氮90%左右,是氮素的主要存在形态。微生物是氮素循环的主要驱动者,在堆肥体系的氮素循环中扮演着极其重要的角色[17]。氮素生理菌群包括固氮菌、氨化细菌、硝化细菌和反硝化细菌等[18]。堆肥中通过添加氮转化和固氮微生物,一方面可以增加堆肥腐熟过程中氮的吸收转化,提高氮素肥效;另一方面,施用到田间或林间,其中的微生物可以长期继续参与环境中氮的转化吸收和利用,起到持效增肥的作用。固氮微生物如固氮菌属(Azotobacter)、根瘤菌属(Rhizobium)等利用固氮酶固定空气中的N2,打开N2分子的三键,生成氨态氮,再通过氨氧化细菌和氨氧化古菌的作用生成亚硝酸盐。硝化细菌如亚硝酸菌属(Nitrosomonas)、硝酸菌属(Nitrobacter)等将亚硝酸盐氧化为硝酸盐等植物可以吸收利用的氮肥。另外,有机质降解时,氨的释放和氨化作用也是细菌参与氮循环增加氮肥效的有效途径。
2.3 其他功能微生物 2.3.1 促生菌微生物菌肥中的促生菌可以调节植物生长和促进植物营养吸收[19]。微生物作用于植物可以产生植物激素,如细胞分裂素、吲哚乙酸等从而调节植物的生长发育和其他生命活动。乙烯是一种天然植物激素,它在植物开花、果熟、衰老和脱落过程中起着重要作用,它对植物次生代谢产物的形成存在抑制和促进双重调控效应,这种调控效应是乙烯与细胞内外多种因子共同作用的结果[20]。菌肥中多种微生物,如芽孢杆菌类细菌可以产生1-羧基-1-氨基环丙烷,降低植物激素乙烯的水平,抑制乙烯对植物的不利影响,保证植物正常生长发育。此外,吲哚乙酸(IAA)也是植物生长素的信号物质,在植物体内普遍存在。研究发现,土壤中约一半以上的细菌可以产生IAA,如固氮螺菌(Azospirillum)、克雷伯氏菌(Kleebsiella)、枯草芽孢杆菌(Bacillus subtilis)等。筛选产生IAA的微生物可为促生生物肥料的研发提供较大帮助。
微生物菌肥可以改善土壤生态,提高土壤中微生物的数量,增加土壤肥力。曹恩晖等[21]通过番茄盆栽试验,向土壤施用复合微生物菌剂,对土壤环境产生的综合效应显著优于空白处理,且经主成分分析得到影响土壤质量的主要土壤环境指标也普遍优于单纯施用化肥、单一功能菌和简单固氮菌组处理的结果。
2.3.2 生防菌菌肥中的有益菌不仅能够促进植物生长、增加作物产量,还能提高植物抗病能力、增强抗逆性。菌肥中放线菌如链霉菌(Streptomyces),可以通过产生抗生素抑制病原菌的生长;木霉属(Trichoderma)真菌可以寄生大丽轮枝孢(Verticillium dahliae),利用微生物进行棉花黄萎病的防治,是目前最有效的方法之一[22];许多微生物与植物存在互生关系,可以诱导植物的系统抗病性,从而抵抗病原菌的入侵;菌肥中的嗜铁细菌能产生嗜铁素吸收土壤中的铁元素形成铁-嗜铁素复合体,进而抑制病原微生物的生长繁殖[23];菌肥还田或还林后,促生菌能在植株根部定殖并利用根际分泌物,抢占病原菌生长位点,使病原菌无法在植株附近生存。
在微生物菌肥中,芽孢杆菌是一类十分重要的细菌。它不仅耐高温对生物质具有极强的降解能力,同时对植物具有显著的生防作用。研究表明,芽孢杆菌BRF21能通过分泌的多种代谢产物对植物病原真菌具有很强的拮抗活性[24];通过基因工程改造的芽孢杆菌,生防效果更加明显。张新建等[25]将外源几丁质酶基因导入巨大芽孢杆菌(Bacillus megat-erium)发现,该工程菌株对病原真菌的拮抗能力明显提高。此外,芽孢杆菌能够分泌一种抑菌蛋白,在酸性或弱碱性条件下具有较强的抗菌活性,但在高温条件下抑菌效果会明显下降。另外,许多微生物能分泌氢氰酸等酶类物质,抑制病原真菌的生长繁殖。
虽然微生物菌肥在生物防治方面有显著作用,但许多问题仍然需要进一步研究。菌肥中许多放线菌分泌次级代谢产物,在杀死病原菌的同时也可能影响促生菌的正常生长,从而影响菌肥对植物的促生作用。另外,菌肥中大量微生物虽然可与植物形成互生或共生关系,但同时也有许多与植物存在竞争关系,影响植物对营养物质的吸收和利用。因此,微生物菌剂接种至堆肥中时,应严格筛选控制菌种,从而保证植物的正常生长。
3 堆肥中微生物菌剂的改良 3.1 微生物菌剂成分的改良纤维素类物质的降解往往需要多种微生物的联合作用,研究表明向堆肥中同时接种含有真菌和细菌的微生物菌剂时,分解纤维素和木质素效率会大大提升,明显高于接种单一菌株的分解腐熟效率,菌肥中添加部分真菌特别是白腐真菌和霉菌,对于木质素的降解具有显著效果[26]。Awasthi等[27]通过分别接种绿色木霉、黑曲霉的真菌孢子悬浮液处理有机废弃物堆肥发现,接种真菌孢子悬浮液并每周搅拌一次的绿色木霉组具有较高的降解率和腐熟度,真菌的添加和堆肥搅拌频率对于有机成分的降解具有显著影响。另外,向堆肥中加入兼性厌氧微生物和嗜热微生物,可有效促进高温缺氧的堆肥内部有机物的降解;添加活性物质可以提高堆肥的发酵效率和腐熟度,减少氮素损耗,增加腐殖质的含量。Liu等[28]向含有鸡粪堆肥的番茄茎秆中分别加入生物炭、泥炭和沸石发现,加入生物炭的实验组堆肥发酵升温时间大大缩短(3 d),最高温度(56℃)明显高于其他实验组(48-51℃)并持续时间较长,加入生物炭的实验组C/N值和挥发性脂肪酸(VFAs)比值都明显高于其他组。Zhang等[29]在堆肥中加入生物炭的同时还加入了菇渣,当加入35%菇渣和20%生物炭时,堆肥的降解率达到最大,生产高质量的堆肥产品的时间由传统的90-270 d缩短至24 d。
但向园林废弃物堆肥中添加其他成分时也应考虑多种因素,如添加的成分是否会影响正常菌种的生长繁殖,抑制发酵反应;是否会增加堆肥腐熟的成本,使投入和产出不平衡。因此,在筛选高效菌种、添加活性物质的同时,也要考虑所增加菌种反应机制研究和堆肥生产的经济效益。
3.2 微生物菌剂施加方式的改造菌肥的施加方式也影响肥效的发挥和经济效益。目前正在研究对植物种子施加丸粒化微生物菌肥,即把菌肥包裹在种子表面,促进种子的萌发和对营养的吸收。在园林废弃物堆肥方面,可以将大体积堆肥分为小体积,使废弃物与菌剂接触面积增大,促进腐熟,但实际应用中还存在着诸多问题,如丸粒化技术成本问题,降解时反应条件如何调节等,需要进一步探究。Nakhshiniev等[30]对园林堆肥进行水热处理,能显著提高茎秆堆肥的稳定性和腐熟度。但是,水热处理后的堆肥材料作为植物栽培基质时,依然可能产生植物性毒素,因此还需要进一步对实验条件等进行研究和处理,从而提高堆肥反应效率。
4 堆肥工艺参数及微生物群落变化监测技术园林废弃物堆肥的降解情况归根结底是功能微生物对有机质的分解程度。通过对堆肥工艺参数及性质变化的检测,以及对功能微生物检测[31],明确堆肥体系的反应进程和微生物群落的变化,以便于有效评估菌肥的腐熟度及肥效。
4.1 目前堆肥化技术中主要检测参数堆肥化技术目前主要从物理、化学、生物学和波谱分析等方面进行检测。
物理检测方面,温度是堆肥过程中的核心参数。作为影响微生物活性的最显著因子,温度对发酵反应速率起着决定性作用,是堆肥体系中微生物生化活动量的宏观指标。一般来讲,堆肥经历升温、维持高温和降温3个过程。通过测量堆肥体系中温度的变化,可以表观的体现腐熟的情况,如堆肥升温过程的特点及堆肥所能达到的最高温度,堆体热量散失过程与温度的关系,控制过程与温度的关系,温度与微生物的生长繁殖及种群演替的关系等。
化学检测方面,在废弃物高温堆肥过程中pH值、C/N值及有机酸含量等变化影响堆肥过程和最终腐熟产品的质量。pH值变化可以比较直观揭示堆肥反应进程,适宜的 pH值可使微生物有效的发挥作用并且保留堆料中有效氮成分,pH值过高或过低都会影响堆肥的效率。C/N值是最常见的判定堆肥腐熟度的评价指标之一,由于堆肥原材料性质的差异,目前大多通过T=(终点C/N比)/(初始C/N比)来评价城市园林废弃物的腐熟度。针对堆肥的不同材料,当T值小于0.7时,可认为堆肥腐熟[32]。有机酸广泛存在于未腐熟堆肥中,随着堆肥的进行,有机酸逐步减少,减少的速度与通气状况和原料相关,即未腐熟的堆肥有机酸含量相对较多,腐熟的堆肥有机酸含量相对较少,可通过研究有机酸的变化评价堆肥腐熟度。
生物学检测方面,在各类堆肥研究中,微生物量的变化大致趋势相同。微生物量均在发酵初期迅速增大,随后大大降低,约一个月之后再次明显上升,堆肥发酵末期时因可代谢营养物的缺乏而微生物生长明显减弱。微生物往往先利用易降解的有机物来维持生长,该有机物殆尽后生长减缓,再转向以较难降解有机物为主要营养物继续生长,至此碳源殆尽生长繁殖再次降低。种子发芽指数GI被认为是最能反映植物毒性大小的腐熟度指标[33],通过检测,当种子发芽指数GI达到50%时,表明堆肥已达腐熟,其植物毒性降至植物能够耐受的水平;当种子发芽指数GI达到80%时,其植物毒性基本消失。
近年来,波谱分析法开始应用,从物质结构角度去评价和分析堆肥过程和腐熟度问题,如红外光谱法和C-核磁共振法[34]。红外光谱法可以辨别化合物的特征官能团,核磁共振法可提供有机分子骨架的信息,能更敏感地反映碳核所处化学环境的细微差别,为测定复杂有机物成分变化提供帮助。但由于堆肥中微生物种类和数量的差异,以及堆肥原料和发酵条件的差异,其有机成分的转化情况存在较大差异,用波谱法测定堆肥腐熟程度,还需要进一步改进。
4.2 功能微生物的监测技术功能微生物是堆肥发酵过程中的关键因素,其种群、数量等动态变化规律可以反应堆肥腐熟变化状况。因此,对于堆肥中功能微生物的鉴定及环境微生物群落的多样性分析显得尤为重要。目前广泛应用的生物技术有DGGE、Roche 454及Illumina MiSeq等[35]。这些技术能较好的揭示堆肥及环境样品中微生物种类,它们之间的相对丰度和变化关系等[36],在探讨不同腐熟阶段微生物多样性以及微生物群落变化、明确功能微生物在堆肥化技术中所起到的作用方面具有重要意义。
DGGE即变性梯度凝胶电泳,是堆肥化技术中微生物群落变化和不同菌群差异检测最广泛的一种手段。其原理是根据样品中DNA浓度的差异在变性剂中解链行为的不同而导致电泳迁移率发生变化,将不同的DNA片段分开[37],在生物多样性调查、基因突变检测等多个领域应用广泛。但该技术往往只能反应出样品中少数优势菌的信息,分辨率和重复性较差、工作量大,应用越来越少。
高通量测序技术为微生物的大量快速检测鉴定提供了较好的平台,MiSeq即是其中较为先进的技术,它以Illumina公司边合成边测序技术为基础,通过专一的可逆终止试剂对数百万个片段同时进行大规模平行测序。该技术具有方便、快捷、成本低、通量高和信息丰富等特点[38]。当每个dNTP加入时,对荧光标记的终止子成像,随后切割,允许下一个碱基的掺入。由于每个测序循环中4种可逆终止子结合的dNTP都存在,所以自然竞争让掺入偏差最小化,根据每个循环的荧光信号测定并检出碱基,与其他技术相比大大降低了原始错误率,实现了可靠的碱基检出。Liu等[39]在“营养添加剂对功能细菌的密度和生物有机肥料的微生物群落结构变化的影响”实验中,通过Illumina-MiSeq测序技术利用16S rDNA库,对实验过程中微生物群落的变化进行分析,在保证测序精确度的同时,大大提高了测序及检测效率。此外,Roche 454测序法、HiSeq 2500技术等都是目前较为先进的测序技术[40],在功能微生物的鉴定,堆肥体系菌群动态变化方面发挥着重要作用。高通量测序技术大大提高了大样本微生物的检测分析效率,全基因组测序技术将会像PCR 技术一样成为微生物学检测鉴定等常规的研究手段[41]。
5 问题与展望目前,我国园林废弃物堆肥产业化发展还处在起步阶段,存在技术成本高、原料来源分散、降解菌种单一及降解机制研究不深入,基础研究脱离生产实践等问题[42]。例如,根瘤菌的固氮作用研究十分深入,有些方面已达到分子水平,但对于植物根际或园林植物残枝上存在的自身固氮菌以及其他功能的促生菌还缺乏深入研究[43];微生物菌剂接种于园林废弃物中的反应条件仍需要进一步探索,接种量与堆肥成本、堆肥效率之间的平衡关系、接种时间以及菌种数量和活性的保持,堆肥生产的标准化和质量检测等方面都需要深入研究。但不容置疑,随着生态园林城市的快速发展,园林绿色废弃物微生物菌剂堆肥处理作为一个新兴的行业,具有广阔的发展前景。国内经济比较发达的城市已经开始对园林植物废弃物进行集中收集,并进行堆肥化处理,如北京、上海、深圳等已颁布了相关政策,鼓励绿色废弃物生产者进行简易堆肥处理,同时免费为其提供堆肥技术,鼓励其建设堆肥厂。
总之,对园林废弃物进行微生物堆肥处理是实现资源的高效循环再利用和可持续发展的重要途径,需要更多的研究者和生产者参与其中,创造更大的经济和社会效益。
| [1] | 田赟. 园林废弃物堆肥化处理及其产品的应用研究[D]. 北京: 北京林业大学, 2012. |
| [2] | 李琬, 刘淼, 张必弦, 等. 植物根际促生菌的研究进展及其应用现状[J]. 中国农学通报, 2014, 30(24): 1-5. |
| [3] | Hassen A, Belguith K, Jedidi N, et al. Microbial characterization during composting of municipal solid waste[J]. Bioresource Technology, 2001, 80(3): 217-225. |
| [4] | 黄丹莲. 堆肥微生物群落演替及木质素降解功能微生物强化堆肥机理研究[D]. 长沙: 湖南大学, 2011. |
| [5] | Paola C, Guido A, Roberto M, et al. Study of the organic matter evolution during municipal solid waste composting aimed at identifying suitable parameters for the evaluation of compost maturity[J]. Waste Management, 2005, 25(2): 209-213. |
| [6] | Boldrin A, Andersen JK, Christensen TH. Environmental assessment of garden waste management in the Municipality of Aarhus, Denmark[J]. Waste Management, 2011, 31(7): 1560-1569. |
| [7] | 隋心, 张荣涛, 钟海秀, 等. 森林生态系统中主要功能微生物的研究进展[J]. 中国农学通报, 2014, 30(28): 1-5. |
| [8] | 张嘉超. 基于分子生物学的堆肥功能微生物种群与体系基质特性关系研究[D]. 长沙: 湖南大学, 2013. |
| [9] | 池玉杰, 伊洪伟. 木材白腐菌分解木质素的酶系统-猛过氧化物酶、漆酶和木质素过氧化物酶催化分解木质素的机制[J]. 菌物学报, 2007, 26(1): 153-160. |
| [10] | Howard RL, Abotsi E, Howard S, et al. Lignocellulose biotechnol-ogy: issues of bioconversion and enzyme production[J]. African Journal of Biotechnology, 2003, 2(12): 602-619. |
| [11] | 史央, 戴传超, 吴耀春, 等. 植物内生真菌强化还田秸杆降解的研究[J]. 环境科学学报, 2004, 24(1): 144-149. |
| [12] | Zeng GM, Huang HL, Huang DL, et al. Effect of inoculating white-rot fungus during different phases on the compost maturity of agricultural wastes[J]. Process Biochemistry, 2009, 44(2): 396-400. |
| [13] | Himmel ME. 生物质抗降解屏障-解构植物细胞壁产生物能[M], 王禄山, 张正, 等译. 北京: 化学工业出版社, 2010. |
| [14] | 曲音波. 木质纤维素降解酶与生物炼制[M], 王禄山, 张正, 等译. 北京: 化学工业出版社, 2011. |
| [15] | DeLaune PB, Moore PA Jr, Daniel TC, et al. Effect of chemical and microbial amendments on ammonia volatilization from composting poultry litter[J]. J Environ Quql, 2004, 33(2): 728-734. |
| [16] | 马丽红, 黄懿梅, 李学章, 等. 牛粪堆肥化中氮素形态与微生物生理群的动态变化和耦合关系[J]. 农业环境科学学报, 2009, 28(12): 2674-2679. |
| [17] | Specht RL. The ratio of foliar nitrogen to foliar phosphorus: a determinant of leaf attributes and height in life-forms of subtropical and tropical plant communities[J]. Australian Journal of Botany, 2010, 58(7): 527-538. |
| [18] | 姚拓, 龙瑞军, 师尚礼, 等. 高寒草地不同扰动生境土壤微生物氮素生理群数量特征研究[J]. 土壤学报, 2007, 44(1): 122-129. |
| [19] | Mantelin S, Touraine B. Plant growth-promoting bacteria and nitrate availability: Impacts on root development and nitrate uptake[J]. Journal of Experimental Biotany, 2004, 55(394): 27-34. |
| [20] | Ruther J, Kleier S. Plant-plant signaling: ethylene synergizes volatile emission in Zea mays induced by exposure to(Z)-3-hexen-1-ol[J]. Journal of Chemical Ecology, 31: 2217-2222. |
| [21] | 曹恩晖, 侯宪文, 李光义, 等. 复合菌剂对盆栽番茄土壤理化性质及微生物活性的影响[J]. 生态环境学报, 2011, 20(5): 875-880. |
| [22] | 王晓杰. 棉花对落叶型黄萎病菌的苗期抗性筛选及绿色木霉菌的生防效果研究[D]. 武汉: 华中农业大学, 2014. |
| [23] | 罗巧玉, 王晓娟, 李媛媛, 等. AM真菌在植物病虫害生物防治中的作用机制[J]. 生态学报, 2013, 33(19): 5997-6005. |
| [24] | 张礼生, 陈红印. 生物防治作用物研发与应用的进展[J]. 中国生物防治学报, 2010, 30(5): 581-586. |
| [25] | 张新建, 黄玉杰, 杨合同, 等. 通过导入几丁质酶基因提高巨大芽孢杆菌的生防效果[J]. 云南植物研究, 2007, 29(6): 666-670. |
| [26] | Lin H, Wang B, Zhunag R, et al. Artificial construction and characterization of a fungal consortium that produces cellulolytic enzyme system with strong wheat straw saccharification[J]. Bioresource Technology, 2011, 102: 10569-10576. |
| [27] | Awasthi MK, Pandey AK, Khan J. Evaluation of thermophilic fungal consortium for organic municipal solid waste composting[J]. Bioresource Technology, 2014, 168: 214-221. |
| [28] | Liu W, Wang ST, Zhang J. Biochar influences the microbial community structure during tomato stalk composting with chicken manure[J]. Bioresource Technology, 2014, 154: 148-154. |
| [29] | Zhang L, Sun XY. Changes in physical, chemical, and microbiological properties during the two-stage co-composting of green waste with spent mushroom compost and biochar[J]. Bioresource Technology, 2014, 171: 274-284. |
| [30] | Nakhshiniev B, Muhammad KB, Hazel BG. Evaluation of hydrothermal treatment in enhancing rice straw compost stability and maturity[J]. Bioresource Technology, 2014, 151: 306-313. |
| [31] | Pan B. Composting of common organic wastes using microbial inoculants[J]. Biotech, 2012, 2(2): 127-134. |
| [32] | 陈同斌, 黄启飞, 高定, 等. 城市污泥堆肥温度动态变化过程及层次效应[J]. 生态学报, 2002, 22(5): 736-741. |
| [33] | Gu W, Zhang F, Xu P, et al. Effects of sulphur and Thiobacillus thioparus on cow manure aerobic composting[J]. Bioresource Technology, 2011, 102: 6529-6535. |
| [34] | 尉良. 农牧业废弃物堆肥腐熟质量控制指标研究[D]. 上海: 东华大学, 2009. |
| [35] | Jennifer AM, Mitchell MH, Kateryna DM, et al. Development and assessment of an optimized next-generation DNA sequencing approach for the mtgenome using the Illumina MiSeq[J]. Forensic Science International: Genetics, 2014, 13: 20-29. |
| [36] | 楼骏, 柳勇, 李延. 高通量测序技术在土壤微生物多样性研究中的研究进展[J]. 中国农学通报, 2014, 30(15): 256-260. |
| [37] | Antonio G, Giovanni C. Compositional shifts of bacterial groups in a solarized and amended soil as determined by denaturing gradient gel electrophoresis[J]. Soil Biology and Biochemistry, 2006, 38(1): 91-102. |
| [38] | Chistoserdova L. Recent progress and new challenges in metageno-mics for biotechnology[J]. Biotechnology Letters, 2010, 32(10): 1351-1359. |
| [39] | Liu L, Li TY, Wei XH, et al. Effects of a nutrient additive on the density of functional bacteria and the microbial community structure of bioorganic fertilizer[J]. Bioresource Technology, 2014, 172: 328-334. |
| [40] | 曹德民, 张穗生, 罗贞贞, 等. 野生型工业酿酒酵母Miseq测序方法的建立[J]. 基因组学与应用生物学, 2014, 33(3): 655-660. |
| [41] | 秦楠, 栗东芳, 杨瑞馥. 高通量测序技术及其在微生物学研究中的应用[J]. 微生物学报, 2011, 51(4): 445-457. |
| [42] | Shaharoona B, Naveed M, Arshad M, et al. Fertilizer-dependent efficiency of Pseudomonas for improving growth, yield, and nutrient use efficiency of wheat(Triticum aestivum L.)[J]. Applied Microbiology and Biotechnology, 2008, 79(1): 147-155. |
| [43] | Liu C, Liu Y, Fan C, et al. The effects of composted pineapple residue return on soil properties and the growth and yield of pineapple[J]. Journal of Soil Science and Plant Nutrition, 2013, 13(2): 433-444. |