




2. 中国农业科学院植物保护研究所 植物病虫害生物学国家重点实验室,北京 100193 ;
3. 中国农业科学院北京畜牧兽医研究所,北京 100193
2. State Key Laboratory for Biology of Plant Diseases and Insect Pests,Institute of Plant Protection,Chinese Academy of Agricultural Sciences,Beijing 100193 ;
3. Institute of Animal Science,Chinese Academy of Agricultural Sciences,Beijing 100193
水稻是我国主要的粮食作物之一,转基因水稻主要有抗虫、抗病、抗除草剂、抗逆环境以及提高水稻品质等多个品种。转cry1Ab/Ac 基因水稻(华恢1 号)是华中农业大学自主培育的对二化螟、三化螟、稻纵卷叶螟等具有较强杀虫活性的水稻品种[1],2009 年转Bt 抗虫水稻华恢1 号和汕优63 获得农业部颁发的农业转基因生物安全证书(生产应用)[2],具有广阔的应用前景。随着全球转基因作物的商业化和大规模种植[3],在一定程度上缓解了粮食紧缺的问题,但转基因作物可能会对动物等非靶标生物或生态环境产生一些非预期效应,所以推广转基因作物的前提是要确保其安全性。
对转基因作物的安全性评价主要从环境和食用安全性两方面考虑,食品安全性评价包括对营养成分、抗营养因子、毒性、过敏性、抗生素抗性等方面进行全面评估[4]。转基因作物被动物摄食后消化规律是否改变,外源基因及蛋白在肠道及粪便中是否残留,是否存在转移到重要器官和血液中的风险;如果动物体内有外源基因和蛋白残留,是否会在人体内积累残留,这些问题都备受关注。大量的研究发现饲喂转Bt 基因或抗除草剂基因的玉米肉鸡,在空肠、回肠、盲肠和排泄物以及重要内脏和血液无外源基因残留[5, 6],在肝脏、鸡蛋排泄物中均未发现外源蛋白的残留[7]。而对转基因水稻中外源基因和蛋白残留的报道仅有两例,发现肉鸡小肠壁、血液、肌肉和脾脏等器官中无cry1Ab 基因残留[8],在肉鸡十二指肠、空肠、回肠内容物和脾脏等器官中无Cry1Ac 蛋白残留[9];迄今国内外尚无转cry1Ab/Ac 基因水稻中外源基因及蛋白在肉鸡体内残留研究的报道。
针对目前基因残留检测中引物单一、覆盖率低等问题,本研究在转基因生物新品种培育科技重大专项的支持下,以饲喂转Bt 基因水稻21 d 和42 d的肉鸡为材料,采用小片段全覆盖的策略,分析肉鸡肠道内容物中cry1Ab/Ac 基因的残留情况,同时采用ELISA 和Western blot 两种方法检测蛋白残留,旨在为转基因水稻安全性评价提供技术支撑和理论依据。
1 材料与方法 1.1 材料 1.1.1 实验材料供试水稻品种为华恢1 号转cry1Ab/Ac 基因水稻,亲本明恢63 为对照组,由华中农业大学提供。
1.1.2 试剂实验所需E. coli DH5α 菌株由抗虫生物技术组保存;pMD19-T 载体等购自TaKaRa 公司;PVDF 膜购自Pall 公司;QIAamp DNA Stool Mini Kit购自QIGEN 公司,QualiPlate 试剂盒购自Envirologix公司。鼠源一抗由本实验室制备,二抗鼠抗羊lgGHRP 购自北京天德悦生物科技公司。
1.2 方法 1.2.1 肉鸡饲喂实验饲养肉鸡日粮配制根据中华人民共和国农业行业标准NY/T33-2004《鸡饲养标准》中肉鸡营养需要。一共140 只鸡分成14 组,其中7 组饲喂含有转基因水稻饲料,另7 组饲喂对照组水稻饲粮。1-21 d 日粮中稻米含量为51.72%,22-42 d 稻米含量为56.54%。
在21 d 和42 d 分别从每个处理中随机取7 只鸡,屠宰前使肉鸡禁食12 h,放血处理后解剖肉鸡,分别取空肠、回肠、盲肠、直肠部位置于离心管中液氮冷冻保存。
1.2.2 肉鸡肠道中外源基因残留检测肉鸡肠道内容物DNA 提取用QIAamp DNA Stool Mini Kit 试剂盒,按照说明书中的方法提取。方法稍作改动,肠道内容物加入1.6 mL Buffer ASL 振荡至均质,加入InhibitEX 药片用于去除杂质和PCR 干扰物,加入蛋白酶K 去除体系中的蛋白,用无水乙醇沉淀DNA,过QIAamp spin 柱洗脱后加入200 μL 超纯水。DNA提取后采用DNA 定量仪(Thermo 公司)测定DNA样品的浓度,并以OD260-280 nm 检测DNA 的纯度。DNA 样品用超纯水稀释至40-50 ng/μL,-20℃储存备用。
为了检测外源基因、水稻内源基因残留,分别根据cry1Ab/Ac 基因以及水稻内源蔗糖磷酸合酶基因SPS(Sucrose phosphate synthase gene,U33175)[10]和淀粉分支酶基因RBE4(Starch branching enzyme,EF055878)[11]、肉鸡内源基因卵清蛋白ov 基因设计引物25 对,其中包括扩增cry1Ab/Ac 基因全长引物1对和100 bp 小片段引物9 对、扩增SPS 基因100 bp小片段引物11 对、扩增RBE4 基因全长引物1 对和100 bp 小片段引物2 对、扩增ov 基因引物1 对,引物序列及扩增长度,见表 1,所有引物由生工生物工程公司合成。

PCR 反应设置水空白对照、阳性对照、待测样品,每个样品做3 个重复。扩增cry1Ab/Ac 基因反应条件:94℃ 预变性5 min ;94℃ 30 s,58℃ 30 s,72℃ 1min 45 s,30 个循环;72℃ 10 min。扩增SPS、ov 基因反应条件:94℃预变性5 min;94℃ 30 s,56℃ 30 s,72℃ 20 s,30 个循环;72℃ 10 min。扩增RBE4 基因反应条件:94℃预变性5 min;94℃ 30 s,54℃ 30 s,72℃ 30 s,30 个循环;72℃ 10 min,PCR 产物4℃保存,1% 琼脂糖凝胶电泳检测。PCR 反应体系,见表 2。

cry1Ab/Ac基因50 ng 和水稻基因组10 μg 加肉鸡肠道内容物5mg 以及对照组cry1Ab/Ac 基因50 ng 和水稻基因组10 μg 分别在37℃温浴0、4、8 和12 h,温浴后的产物用QIAamp DNA Stool Mini Kit 试剂盒提取DNA,DNA 产物用根据cry1Ab/Ac 基因所设计的10 对引物检测cry1Ab/Ac 基因片段。PCR 反应条件同1.2.2,1%琼脂糖凝胶电泳检测。
1.2.4 肉鸡肠道中的外源蛋白残留检测肉鸡肠道内容物总蛋白用PBS(pH7.4)缓冲液提取,总蛋白定量采用BCA 方法,试剂盒购自北京艾德莱生物科技有限公司。Cry1Ab/Ac 蛋白残留检测采用ELISA双抗夹心法,QualiPlate 试剂盒购自美国Envirologix公司,方法按照试剂盒说明书进行,重复3 次;其中放置温度均改为37℃,检测结果用Western blot 验证。Western blot 方法如下:待测样经过SDS-PAGE分离后转印至PVDF 膜上,将膜用TBS-T 溶液漂洗后用20 mL 5% 脱脂奶粉封闭2 h,加入含一抗(0.5μL)的TBS-T 漂洗液15 mL,4℃孵育过夜或室温1h,加入含鼠抗兔IgG 二抗(1 μL)的TBS-T 漂洗液15 mL,室温1 h,每步结束后漂洗膜3 次,每次10min,最后膜用显色液显色后拍照。
2 结果 2.1 肉鸡肠道中外源基因残留检测肉鸡肠道内容物经QIAamp DNA Stool MiniKit 试剂盒提取后,DNA 定量仪检测DNA 的纯度(OD260-280 nm)均在1.7-1.9,将DNA 样品用超纯水稀释至40-50 ng/μL,每个样品取1.0 μL 用于检测。PCR 方法分别检测了肉鸡肠道内容物提取的总DNA中cry1Ab/Ac 基因全长和9 个小片段基因,琼脂糖凝胶电泳结果显示,仅能扩增出10 个片段的相应正对照,在肠道内容物总DNA 中,均未检测到 基因全长和9 个小片段(图 1-A 和图 1-B)。同样,在空肠、回肠、盲肠和直肠内容物提取的总DNA 中,也检测不到水稻内源基因SPS 及REB4(图 1-C、图 1-D和图 1-E);但所有的内容物总DNA 中都可以检测到肉鸡内源ov 基因(图 1-F)。

|
| 图 1 肉鸡肠道内容物提取的总DNA 中cry1Ab/Ac 基因的 PCR 检测结果 A :cry1Ab/Ac 基因全长(1 830 bp)检测;B :cry1Ab/Ac 基因片段(100 bp)检测;C :水稻中内源SPS 基因片段(100 bp)检测;D :水稻中内源RBE4全长(484 bp)检测。E :水稻中内源RBE4 片段(100 bp)检测;F :肉鸡内源ov 基因(396 bp)检测。M:Marker(A:DL2000;B、D、E:DL500;C、F :Marker Ⅰ);- :超纯水;+ :华恢1 号米粉基因组(F 组为肉鸡肠道内容物);K :空肠;H :回肠;M :盲肠;Z :直肠 |
从消化时间上看,不加肠道内容物对照处理中12 h 仍能扩增出全长cry1Ab/Ac 基因;然而,cry1Ab/Ac 基因在加入了肠道内容物后,在4 h 就检测不到全长cry1Ab/Ac 基因(图 2-A);但是在4 和8 h 时还能检测出cry1Ab/Ac 基因9 个小片段,而到达12 h 时,9 个小片段均无法检测到(图 2-B)。

|
| 图 2 体外消化cry1Ab/Ac 基因全长(A)及片段(B) M1 :BM2000 ;M2 :DL500 DNA Marker ;1 :0 h 总DNA ;2 :0 h 总DNA 加内容物;3 :4 h 总DNA 加内容物;4 :8 h 总DNA 加内容物;5 :12 h 总 DNA 加内容物;6 :12 h 总DNA |
ELISA 检测前为避免样品中的蛋白浓度对检测结果产生较大影响,先用BCA 法对提取的蛋白进行定量,定量至20 μg。用双抗夹心法对样品进行检测,检测结果如表 3 所示,检测结果中21 d、42d 转基因和非转基因组中盲肠和直肠样品21CM、21CZ、21TM、21TZ、42CM、42CZ、42TM 和42TZ出现比较明显的显色反应,检测值表明其中可能存在Cry1Ab/Ac 蛋白残留。
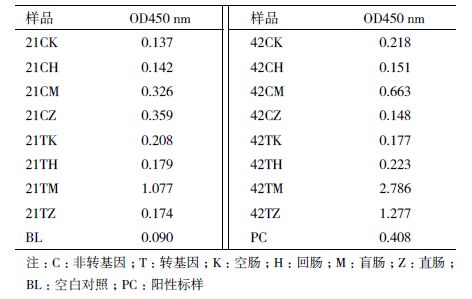
对ELISA 检测结果中产生颜色反应的样品进行SDS-PAGE 和Western blot 分析,结果显示SDSPAGE电泳图中在60 kD 处42CZ 样品和42TZ 样品存在疑似条带,但Western blot 分析结果表明这两个样品中并不存在Cry1Ab/Ac 蛋白。
3 讨论图 3家禽肠道中没有Bt 蛋白的结合位点,因此,Bt蛋白不会破坏家禽的肠道继而影响家禽发育。但如果转Bt 基因水稻中的外源蛋白不易消化,蛋白残留可能会对家禽产生致敏性[12],基因残留也可能引起其他非预期效应,需要对转Bt 基因水稻的消化情况进行分析。本研究发现饲喂转Bt 水稻肉鸡肠道中没有cry1Ab/Ac 基因和蛋白残留,同时用体外消化实验进行了进一步的验证。
针对基因残留检测中引物单一、覆盖率低等问题,本研究采用多对引物同时对cry1Ab/Ac 基因全长及基因碎片进行检测。在饥饿12 h 的肉鸡肠道内容物中未检测到cry1Ab/Ac 基因的残留,这与Rossi 和Tony 的结果是一致的。Rossi[13]在饲喂转Bt 玉米肉鸡肠道中发现外源基因仅在肉鸡肠道消化系统的前面部分嗉囊中检测到,原因是肉鸡在这一时间段不可能完全消化掉饲料。本研究中水稻内源基因SPS和REB4 与cry1Ab/Ac 基因有同样的降解情况,而Tony 等[14]在肉鸡肠道空肠之后的部位能检测到植物内源基因的残留。为了进一步验证动物实验中外源基因在肉鸡肠道中的降解情况,本研究进行了体外模拟实验,结果表明全长cry1Ab/Ac 基因在4 h 已经降解,而小片段在12 h 也降解完全,验证了在饥饿12 h 的肉鸡肠道内容物中检测不到cry1Ab/Ac 基因的结果。

|
| 图 3 21 d、42 d 转基因和非转基因饲喂肉鸡盲肠和直肠内容物总蛋白的SDS-PAGE 分析(A)及Western blot分析(B) M :蛋白质Marker ;1 :Cry1Ab/Ac 蛋白;2 :21CM ;3 :21CZ ;4 :21TM ;5 :21TZ ;6 :42CM ;7 :42CZ ;8 :42TM ;9 :42TZ。其中,C :非转基因;T :转基因;K :空肠;H :回肠;M :盲肠;Z :直肠 英文标题 |
本研究ELISA 实验中一些非转基因样品也出现了明显颜色反应,这与Chowdhμry[15] 和Einspanier[16]的结果相类似,肉鸡肠道内容物中一些外源或内源的污染物可能会引起显色反应,影响实验的结果。Western blot 检测发现样品中并没有Bt蛋白残留[17],与本研究结果一致。随着肠道消化吸收食物,在肠道以外部分如鸡的蛋[18]、血液[5]、肌肉[19, 20]、内脏组织[6, 21]、粪便[7]中也未检测到外源蛋白的残留。
上述研究充分说明转基因水稻中的外源基因和蛋白在肉鸡肠道内容物中无残留,本实验室进一步利用宏基因组16S rDNA 测序技术发现饲喂转Bt 基因水稻对肉鸡肠道微生物群落结构无显著影响(数据待发表),当然还需要营养学、毒理学等方面的研究对转Bt 基因水稻食用安全性做出全面的评价。
4 结论本研究分析了饲喂转Bt 水稻肉鸡肠道中外源基因和蛋白的残留情况,结果显示,肉鸡肠道内容物中没有cry1Ab/Ac 基因和蛋白残留,该研究体系为其他转基因作物对动物饲喂安全性评价提供了借鉴。
| [1] | Tu JM, Zhang GA, Datta K, et al. Field performance of transgeniclite commercial hybrid rice expressing Bacillus thuringiensis deltaendotoxin[J]. Nature Biotechnology, 2000, 18(10): 1101-1104. |
| [2] | Lu C. The first approved transgenic rice in China[J]. GM Crops,010, 1(3): 113-115. |
| [3] | James C. Global Status of Commercialized Biotech/GM Crops : 013[R]. ISAAA Brief 46, 2014. |
| [4] | 许文涛, 贺晓云, 黄昆仑, 等. 转基因植物的食品安全性问题及价策略[J]. 生命科学, 2011, 23(2): 179-185. |
| [5] | Swiatkiewicz S, Koreleski J, Arczewska-Wlosek A, et al. Detectionf transgenic dna from Bt maize and herbicide tolerant soybean mealn tissues, eggs and digestive tract content of laying hens fed dietsontaining[J]. Annals of Animal Science, 2011, 11(3): 413-24. |
| [6] | Jennings JC, Albee LD, Kolwyck DC, et al. Attempts to detectransgenic and endogenous plant DNA and transgenic protein inuscle from broilers fed Yield Gard Corn Borer Corn[J]. Poultrycience, 2003, 82(3): 371-380. |
| [7] | Ash J, Novak C, Scheideler SE. The fate of genetically modifiedrotein from Roundup Ready Soybeans in laying hens[J]. Journalf Applied Poultry Research, 2003, 12(2): 242-245. |
| [8] | 杜红方. 转基因水稻作为肉仔鸡日粮原料的安全性评价[D].京: 中国农业科学院, 2006. |
| [9] | 秦海峰. 转 Cry1Ac/sck 基因糙米作为肉仔鸡日粮原料的安全性价[D]. 北京: 中国农业科学院, 2012. |
| [10] | Ding JY, Jia JW, Yang LT, et al. Validation of a rice specific gene,ucrose phosphate synthase, used as the endogenous referenceene for qualitative and real-time quantitative PCR detection ofransgenes[J]. Journal of Agricultural and Food Chemistry,2004, 52(11): 3372-3377. |
| [11] | Jeong SC, Pack IS, Cho EY, et al. Molecular analysis anduantitative detection of a transgenic rice line expressing aifunctional fusion TPSP[J]. Food Control, 2007, 18(11): 434-1442. |
| [12] | Bannon GA. What makes a food protein an allergen?[J]. Currentllergy and Asthma Reports, 2004, 4(1): 43-46. |
| [13] | Rossi F, Morlacchini M, Fusconi G, et al. Effect of Bt corn onroiler growth performance and fate of feed-derived DNA in theigestive tract[J]. Poultry Science, 2005, 84(7): 1022-1030. |
| [14] | Tony MA, Butschke A, Broll H, et al. Safety assessment of Bt 176aize in broiler nutrition : Degradation of maize-DNA and itsetabolic fate[J]. Archives of Animal Nutrition-Archiv Furierernahrung, 2003, 57(4): 235-252. |
| [15] | Chowdhury EH, Shimada N, Murata H, et al. Detection of Cry1Abrotein in gastrointestinal contents but not visceral organs ofenetically modified Bt11-fed calves[J]. Vet Hum Toxicol, 2003,5(2): 72-75. |
| [16] | Einspanier R, Lutz B, Rief S, et al. Tracing residual recombinanteed molecules during digestion and rumen bacterial diversity inattle fed transgene maize[J]. European Food Research andechnology, 2004, 218(3): 269-273. |
| [17] | Lutz B, Wiedemann S, Einspanier R, et al. Degradation of Cry1Ab protein from genetically modified maize in the bovineastrointestinal tract[J]. J Agric Food Chem, 2005, 53(5): 453-1456. |
| [18] | Ma QG, Gao CQ, Zhang JY, et al. Detection of transgenic andndogenous plant DNA fragments and proteins in the digesta,lood, tissues, and eggs of laying hens fed with phytase transgenicorn[J]. PLoS One, 2013, 8(4): 10. |
| [19] | Gao CQ, Ma QG, Zhao LH, et al. Effect of dietary phytaseransgenic corn on physiological characteristics and the fate ofecombinant plant DNA in laying hens[J]. Asian-Australasianournal of Animal Sciences, 2014, 27(1): 77-82. |
| [20] | Stadnik J, Karwowska M, Dolatowski ZJ, et al. Effect of geneticallyodified, insect resistant corn(Mon 810)and glyphosate tolerantoybean meal(Roundup Ready)on physico-chemical propertiesf broilers’ breast and thigh muscles[J]. Bulletin of theeterinary Institute in Pulawy, 2011, 55(3): 541-546. |
| [21] | Sieradzki Z, Mazur M, Kwiatek K, et al. Assessing the possibilityf genetically modified DNA transfer from GM feed to broiler,aying hen, pig and calf tissues[J]. Polish Journal of Veterinaryciences, 2013, 16(3): 435-441. |