




2.甘肃省农业科学院作物研究所,兰州 730070;
3. 中国农业科学院生物技术研究所,北京 100081
2. Institute of Industrial Crops,Gansu Academy of Agricultural Sciences,Lanzhou 730070;
3. Biotechnology Research Institute,Chinese Academy of Agricultural Sciences,Beijing 100081
胡麻,即油用型亚麻的俗称,是世界古老的油料和纤维作物之一,其栽培历史已有8 000多年[1],主要分布在中国、加拿大、美国、印度、欧洲、非洲、南美等国家和地区[2]。在我国,胡麻也有2 000多年的栽培历史[3],主要分布在甘肃、山西、内蒙、宁夏、河北、新疆、青海等省区,年种植面积约33万hm2左右[4]。胡麻籽油富含omega-3脂肪酸,以多不饱和脂肪酸α-亚麻酸、油酸、亚油酸为主要成分,其中α-亚麻酸含量高达50%以上,对人体健康十分有益,具有预防心脑血管疾病、提高儿童免疫力、促进大脑发育、降血脂、稳定血糖、减肥等多种功能[5]。作为重要的人体有益脂肪酸来源作物,胡麻籽的营养生理生化研究已成为国内外的研究热点。对胡麻品质育种而言,脂肪酸遗传机制研究及高油高亚麻酸品种创新更显迫切。
胡麻脂肪酸含量是典型的数量性状,有关其遗传规律的研究较少。重组自交系(Recombinant inbred lines,RIL)是作物数量性状遗传分析的理想群体材料,目前广泛应用于各种作物数量性状的遗传分析、图谱构建与QTL定位等研究[6, 7, 8]。利用RIL群体进行数量性状遗传分析的主要优点是,它是一种永久性群体,可以进行有重复的比较试验,适合于环境影响较大的复杂性状的遗传研究[9]。应用RIL群体进行胡麻数量遗传与作图研究并不多见,仅Cloutier等[10]利用3个RILs群体构建了含有15个连锁群的亚麻遗传图谱。随着数量性状遗传研究的深入,构建一套可供重复利用的胡麻重组自交系材料十分必要,可为相关研究奠定基础。为此,本研究以国内广泛种植的胡麻品种陇亚8号和国外引进纤用品种阿里安为亲本组合,构建一套含有162份家系的RIL群体。该群体农艺性状变异丰富,但品质状况尚待分析。为更好地利用该RIL群体,采用气相色谱法测定该群体的脂肪酸含量,对其遗传变异与遗传机制等开展研究,同时筛选优异株系,旨在为胡麻品质改良提供新材料。
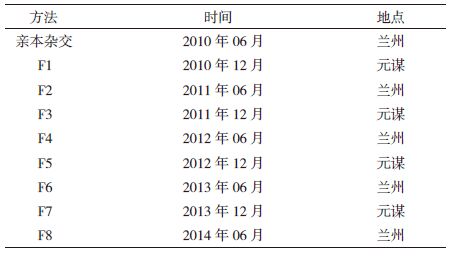
1 材料与方法 1.1 材料以亲本组合“陇亚8号×阿里安”构建的胡麻重组自交系(RIL)F2:8群体为实验材料,构建程序如图 1所示,通过逐代进行单株选择而成,共包括162份家系材料。其亲本陇亚8号为国内育成的油用型主栽胡麻品种,主要特点是高产稳产、优质抗病,阿里安为国外引进的纤维型亚麻品种。两亲本及RIL群体之间农艺性状差异显著,变异广泛,但其品质性状的变异及遗传有待进一步分析评价。

|
| 图 1 脂肪酸含量在重组自交系中的频次分布图 |
田间试验地点设在甘肃省景泰县畜牧业良种场,位于东经104°04´、北纬37°10´,海拔1 616 m,属沿黄灌区。亲本及RIL群体材料于2014年在甘肃省景泰县畜牧业良种场种植,采用顺序排列,单行区种植,行长1 m,每行播种100粒,3次重复。成熟后每行随机取10株混样脱粒,进行品质测定与分析。
1.2.2 粗脂肪测定采用Hanon SOX406型脂肪测定仪进行。脂肪酸含量测定利用气相色谱法在Agilent 7820A GC型气相色谱分析系统上完成。其GC条件为:进样口温度250℃,FID温度:300℃,分流比60:1;柱流速(N2):2 mL/min,恒流模式:H2流速:40 mL/min,空气流速:400 mL/min,尾吹气(N2)流速:30 mL/min;柱温程序:210℃恒温8 min,色谱柱(AT-FFAP):30 m×320 mm×0.33 μm;采用峰面积归一法对脂肪酸进行定量。
1.2.3 数据处理与分析应用Excel和DPS软件进行数据处理与相关统计分析。脂肪酸含量的遗传分析采用盖钧镒等[11]提出的主基因+多基因混合遗传模型中的单个分离世代数量性状分离分析方法,分析软件由南京农业大学章元明教授提供[12]。
2 结果 2.1 亲本及RIL群体脂肪酸含量变异亲本及RIL群体粗脂肪和脂肪酸含量列于表 2。亲本陇亚8号为国内育成油用型胡麻品种,粗脂肪含量38.52%,脂肪酸含量分别为亚麻酸44.42%、油酸31.00%、亚油酸12.66%、棕榈酸5.75%、硬脂酸5.54%。阿里安为纤用型亚麻品种,粗脂肪含量35.44%,脂肪酸含量分别为亚麻酸49.05%、油酸29.05%、亚油酸12.42%、棕榈酸5.58%、硬脂酸5.23%。两亲本之间粗脂肪和亚麻酸含量差异较大,分别相差3.08%和4.63%,其它4种脂肪酸含量差异不大。由二者杂交衍生而成的重组自交系(RIL)粗脂肪含量变幅为35.68%-41.46%,变异系数3.36%,最小值接近低亲阿里安,最大值为41.46%,较高亲陇亚8号高2.94%。5种脂肪酸组分中,硬脂酸含量变异最大,变异系数11.89%,变幅为3.24%-6.29%;其次为油酸含量,变异系数7.58%,变幅为20.44%-30.73%;其它3种脂肪酸变异系数在4%-5%,其中亚麻酸含量的变幅较大,最低含量为45.16%,最高含量达57.37%,属高亚麻酸含量株系材料。总体来看,RIL群体家系材料间粗脂肪和脂肪酸含量亦存在较为广泛的变异,表现超亲分离现象,有必要开展进一步遗传分析。

RIL家系粗脂肪和脂肪酸含量的分布参数偏度值和峰度值(表 2)显示,其偏度和峰度绝对值均小于1,表明粗脂肪和脂肪酸含量的分布近似呈正态分布,属多基因控制的数量性状。为直观的揭示其分布特征,分别以粗脂肪和脂肪酸含量为横坐标,分布次数为纵坐标,绘制二维次数分布图。结果(图 1)显示,RIL群体粗脂肪和脂肪酸含量均呈连续变异,表现出数量性状的典型分布特征,并且多呈现偏态或多峰分布现象,可能存在主基因效应。
2.3 脂肪酸含量的遗传分析利用目前广泛应用的数量性状主基因+多基因混合遗传模型中的单个世代分离分析方法,对RIL群体粗脂肪和脂肪酸含量进行遗传分析。通过极大似然法和IECM 算法估算各种模型的分布参数,根据AIC准则和适合性检验选取最佳遗传模型,即AIC值较小且适合性检验统计量达到显著水平数量最少的模型为最优遗传模型,并在最优模型下估算相应的遗传参数。通过分析软件运算,得到各性状不同遗传模型的极大似然函数、AIC值及适合性检验统计参数。结果(表 3)表明,备选遗传模型的所有统计量均未达到显著水平,按照AIC最小准则,筛选得到各性状的最优遗传模型及遗传参数。

初步遗传分析结果(表 3)表明,粗脂肪含量的最优遗传模型为3MG-CEA,即粗脂肪含量受3对等加性主基因控制,主基因遗传率为85%;5种脂肪酸组成中,亚麻酸含量的最优遗传模型为2MG-Duplicate,即亚麻酸含量受2对重叠作用主基因控制,主基因遗传率为36%;亚油酸含量的最优遗传模型为3MG-CEA,即亚油酸含量受3对等加性主基因控制,主基因遗传率为80%;油酸、棕榈酸和硬脂酸含量的最优遗传模型均为0MG,即均表现为无主基因效应,其遗传可能受微效多基因控制。
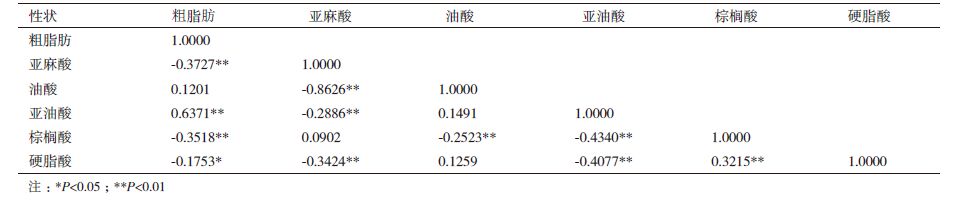
2.4 粗脂肪与脂肪酸组分的相关分析RIL群体粗脂肪与脂肪酸组分之间的相关系数结果(表 4)表明,粗脂肪与亚油酸含量存在显著的正相关,与亚麻酸、棕榈酸、硬脂酸存在显著的负相关;5种脂肪酸组分之间,亚麻酸含量与油酸、亚油酸及硬脂酸存在显著的负相关,这与前人分析结果基本一致[13, 14]。由粗脂肪与脂肪酸组分之间的相关性说明,在RIL群体中可能筛选出一些高油高亚油酸、高亚麻酸的优良家系材料,作为创新资源在胡麻品质改良中加以利用。
由前面分析可知,RIL家系粗脂肪和脂肪酸含量存在广泛变异,多呈超亲分离现象。粗脂肪与脂肪酸含量的超亲优势(表 5)显示,162份家系的粗脂肪含量均高于低亲,超低亲优势在0.68%-16.99%;其中粗脂肪含量高于高亲的家系有64份,超高亲比例为39.5%,超高亲优势在0.10%-7.63%。162份家系的亚麻酸含量也均高于低亲,超低亲优势在1.67%-29.15%;其中亚麻酸含量高于高亲的家系148份,超高亲比例达91.4%,超高亲优势0.04%-16.96%,可见亚麻酸含量的超高亲优势十分明显。亚油酸含量有96份材料高于低亲,其中有77份材料高于高亲,超高亲比例为47.5%,超高亲优势在0.08%-11.06%。油酸含量除2份家系材料介于双亲之间外均低于低亲,棕榈酸和硬脂酸也均存在双向超亲优势。可以看出,RIL群体粗脂肪、亚麻酸、亚油酸含量的超高亲优势普遍存在,由此可以筛选出高油高亚油酸家系材料5份,高亚麻酸家系材料6份,作为优异种质新材料在胡麻品质育种中进行创新应用。

采用气相色谱法对由陇亚8号/阿里安亲本组合衍生的重组自交系的品质状况进行了准确分析,结果表明,该RIL群体粗脂肪和脂肪酸组成亦存在广泛变异,表现超亲分离现象,分布近似为正态分布,呈现数量性状的典型分布特征,属多基因控制的数量性状。有关数量性状遗传机制的理论与研究方法,目前广泛应用的主要是盖钧镒等[11]提出的主基因+多基因理论体系。该理论认为,控制数量性状的基因数目有多有少,各对基因效应大小不等且易受到环境影响。将效应大的、在一般条件下可以检测出来的基因称为主基因;效应小的、在现有试验条件下即使通过专门技术仍然检测不出来的基因称之为多基因。RIL群体是遗传上稳定的永久分离群体,可以进行重复实验,进而减少环境影响,提高数量性状基因检测效率和遗传效应估计精度。章元明等[15]相继建立了利用RIL 群体进行数量性状遗传体系分析的方法。本研究初次运用该方法,利用胡麻RIL群体对粗脂肪和脂肪酸含量遗传规律进行初步分析,结果表明粗脂肪含量为3对主基因遗传,主基因遗传率为85%;5种脂肪酸组成中,亚麻酸含量为2对重叠作用主基因遗传,主基因遗传率为36%;亚油酸含量为3对主基因遗传,主基因遗传率为80%;油酸、棕榈酸和硬脂酸含量均表现为无主基因效应,属多基因遗传。以上分析虽然对主基因效应作出了初步遗传估计,但由于未加入不分离世代P1、P2,因此无法对多基因效应作出准确鉴别。大豆种子脂肪酸含量的遗传分析表明[16],5种脂肪酸遗传主要受主基因控制,这与本研究结果有所不同。胡麻粗脂肪与亚麻酸、亚油酸为主基因遗传,而油酸、棕榈酸和硬脂酸遗传主要受多基因控制。
3.2 胡麻品质育种的探讨胡麻是十分优质的油料作物之一,胡麻籽粗脂肪含量一般在38%-40%,胡麻油脂中脂肪酸构成以对人体有益的omega-3不饱和脂肪酸为主,其中亚麻酸含量高达50%-55%,油酸和亚油酸含量占30%-40%,具有很高的营养保健价值。随着我国胡麻产业的发展和人们生活水平的提高,胡麻育种与品质改良显得日益重要。胡麻高值化技术已经作为我国胡麻现代产业技术体系的重点任务进行研发,而胡麻品质改良与高值化技术的核心就是提高粗脂肪和亚麻酸含量。本研究通过对胡麻重组自交系粗脂肪和脂肪酸含量的分析发现,RIL群体的粗脂肪和脂肪酸含量存在广泛变异和超亲分离现象,超亲优势明显,粗脂肪、亚麻酸和亚油酸含量的超高亲优势分别达7.63%、16.96%和11.06%。因此,通过常规重组自交技术培育优异品系是改良胡麻品质的有效方法。相关分析表明,RIL群体粗脂肪与亚油酸含量存在显著正相关,与亚麻酸含量存在显著负相关,利用RIL群体可能筛选出一些高油高亚油酸或高亚麻酸含量的优异品系材料。本研究筛选出高油高亚油酸和高亚麻酸含量的优良稳定株系11份,可供胡麻品质育种进一步鉴定利用。
4 结论胡麻RIL群体的粗脂肪与脂肪酸含量存在广泛变异,表现超亲分离现象,其分布近似为正态分布,呈现数量性状连续变异的典型分布特征。通过运用主基因+多基因遗传模型分析表明,粗脂肪含量为3对等加性主基因遗传,主基因遗传率为85%;5种脂肪酸组成中,亚麻酸含量为2对重叠作用主基因遗传,主基因遗传率为36%;亚油酸含量为3对等加性主基因遗传,主基因遗传率为80%;油酸、棕榈酸和硬脂酸含量均表现为无主基因效应的多基因遗传。同时筛选出高油高亚油酸和高亚麻酸含量的优良稳定株系11份。
| [1] | Sabine K. New research on the cultural history of the useful plant Linum usitatissimum L. (flax), a resource for food and textiles for 8 000 years[J]. Veget Hist Archaeobot, 2011, 20:507-508. |
| [2] | Sirisha M, Shyam Sundar R, Amareshwar P. Know more about propitious seed-flax[J]. International Journal of Pharmacy & Technology, 2010, 2:246-258. |
| [3] | Liu FH, Chen X, Long B, et al. Historical and botanical evidence of distribution, cultivation and utilization of Linum usitatissimum L. (flax)in China[J]. Veget Hist Archaeobot, 2011, 20:561-566. |
| [4] | 王利民. 我国胡麻生产现状及发展建议[J]. 甘肃农业科技, 2014, 4:60-61. |
| [5] | Nirmala Halligudi. Pharmacological properties of Flax seeds:A Review[J]. Hygeia J D Med, 2012, 4(2):70-77. |
| [6] | 李法计, 常鑫, 王宇娟, 等. 小麦重组自交系群体9个重要农艺性状的遗传分析[J]. 麦类作物学报, 2013, 33(1):23-28. |
| [7] | 陈四龙, 程增书, 王瑾, 等. 花生远缘杂交重组系的遗传变异与优良品系综合选择[J]. 中国油料作物学报, 2013, 10:156-163. |
| [8] | 侯萌, 齐照明, 韩雪, 等. 大豆蛋白质和油分含量QTL定位及互作分析[J]. 中国农业科学, 2014, 47(13):2680-2689. |
| [9] | 王金社, 李海旺, 赵团结, 等. 重组自交家系群体4 对主基因加多基因混合遗传模型分离分析方法的建立[J]. 作物学报, 2010, 36(2):191-201. |
| [10] | Cloutier S, Ragupathy R, Miranda E, et al. Integrated consensus genetic and physical maps of flax(Linum usitatissimum L.)[J]. Theor Appl Genet, 2012, 125:1783-1795. |
| [11] | 盖钧镒, 章元明, 王建康. 植物数量性状遗传体系[M]. 北京:科学出版社, 2003. |
| [12] | 曹锡文, 刘兵, 章元明. 植物数量性状分离分析Windows软件包SEA 的研制[J]. 南京农业大学学报, 2013, 36(6):1-6. |
| [13] | 赵利, 党占海, 张建平, 等. 不同类型胡麻品种资源品质特性及其相关性研究[J]. 干旱地区农业研究, 2008, 26(5):6-9. |
| [14] | 赵利, 党占海, 张建平. 甘肃胡麻地方品种种质资源品质分析[J]. 中国油料作物学报, 2006, 28(3):282-286. |
| [15] | 王金社, 赵团结, 盖钧镒. 回交自交系(BIL)群体4 对主基因加多基因混合遗传模型分离分析方法的建立[J]. 作物学报, 2013, 39(2):198-206. |
| [16] | 李侠, 常玮, 韩英鹏, 等. 大豆种子脂肪酸含量的遗传分析[J]. 大豆科学, 2009, 28(3):403-408. |