




2. 河南新乡华星药厂,新乡 453007;
3.河南省一科技部共建细胞分化国家重点实验室培育基地和河南省生物工程重点实验室,新乡 453007
2. Henan Xinxiang Hua Xing Pharmaceutical Factory,Xinxiang 453007;
3. State Key Laboratory Cultivation Base for Cell Differentiation Regulation and Henan Bioengineering Key Laboratory,Henan Normal University,Xinxiang 453007
豹蛙抗瘤酶(Onconase,ONC),又称豹蛙酶,是从北极豹蛙卵母细胞和早期胚胎中提取的一种天然蛋白,是核糖核酸酶A家族的最小成员[1],是第一个进入抗肿瘤临床试验的核糖核酸酶[2, 3],在体内外多种肿瘤试验中体现出了很强的杀伤作用。1996年以来,Tamir公司(前身为Alfacell公司)针对ONC已开展乳腺癌、胰腺癌、非小细胞肺癌、恶性间皮质瘤等的Ⅰ-Ⅲ期临床试验,均表现出了良好的抗肿瘤作用[4]。ONC是一个由104个氨基酸残基构成的单链蛋白,含4对二硫键。与RNaseA相比,尽管属同一超家族成员,并且氨基酸序列具有30%相似性,但在酶活和细胞毒性上具有成千上万倍的差别。RNA降解活性为ONC的1 000倍,而ONC的细胞毒性达到RNaseA的10 000倍[5]。ONC较高的细胞毒性和较低的免疫原性正是它成为肿瘤药物的关键因素。ONC的作用机制尚不完全明确,目前认为它通过静电作用与细胞膜表面结合,再通过膜流和内吞作用进入细胞内[6]。在胞内通过特异性降解tRNA而抑制蛋白质合成[7],Qiao等[8]研究发现ONC特异性降解miRNA前体为ONC的作用机提出新的研究思路。
目前,临床试验中所用的ONC都取自蛙卵和早期胚胎,资源少、周期长、收率低及代价高,难以满足临床需要[9]。ONC常见的已表达系统有大肠杆菌系统和毕赤酵母系统,但大肠杆菌系统需要进行包涵体复性等繁琐步骤,活性难以保证,不利于工业化生产[9, 10, 11];毕赤酵母系统因表达量过低(150 mg/L)[12],不利于大规模应用。与之相反,HSA的毕赤酵母表达技术已经十分成熟,表达量高(最高表达量10 g/L)[13, 14]且分离纯化体系完善,实用优势明显,而且人血清白蛋白有重要的生物学作用和广泛的临床应用,常用来与其他蛋白融合以延长蛋白半衰期,是理想的药物载体[15]。
本研究拟利用巴斯德毕赤酵母表达人血清白蛋白和豹蛙抗瘤酶的融合蛋白,将rHSA的高表达和纯化成熟的优势体现于ONC,并对其表达条件和分离纯化方法进行优化,旨在有效降低ONC的生产成本,为其大规模生产应用提供参考。。
1 材料与方法 1.1 材料 1.1.1 菌株、质粒和基因大肠杆菌(Escherichia coli)DH5α菌株购自武汉晶赛生物工程技术有限公司,毕赤酵母X-33、GS115、SMD1168菌株,分泌型酵母表达载体pPIC9、pPIC9K、pPICZα-A购自Invitrogen公司,HSA-ONC融合基因委托上海生工生物工程有限公司合成。人肝癌细胞株7402、HeLa、RH35细胞株菌购自中国医学科学院基础医学研究所细胞资源中心(北京)。
1.1.2 试剂和工具酶DMEM、RPMI-1640培养基、青霉素、链霉素购自Invitrogen公司,胎牛血清购自杭州天杭生物科技有限公司,MTT、DMSO购自Geneview公司。DNA片段回收试剂盒及质粒快速提取试剂盒均购自鼎国生物技术有限公司,酵母基因组快速提取试剂盒购自北京博迈德科技发展有限公司。限制性内切酶Pme I、Bst B I、Bam H I、Eco R I购自MBI公司,Protein Ladder购自上海赛默飞世尔科技有限公司。DEAE Sepharose Fast Flow购自美国GE公司。HSA一抗购自北京中杉金桥生物技术有限公司,ONC一抗购自武汉博士德生物工程有限公司,其他试剂为国产分析纯。
1.1.3 培养基及基础培养条件常用培养基:LB、YPD、BMGY、PTM1、PTM2等培养基参考Invitro-gen公司的酵母表达手册。
BSM(Basic Salt Medium,BSM)(g/L):硫酸钾14.3,硫酸镁 11.7,盐酸胍6,氢氧化钾3.9,硫酸钙0.9,甘油40,磷酸23.5 mL/L。
LBSM(Low Salt Basic Salt Medium,LSBSM)(g/L):硫酸钾4.55,硫酸镁3.73,氢氧化钾1.03,硫酸钙0.23,六偏磷酸钠6.5,甘油40。
摇瓶培养条件:温度28℃,培养转速300 r/min,诱导转速200 r/min,诱导10 d。发酵罐发酵条件:发酵开始至12 h:温度28℃,转速400 r/min,通气量0.2 NM3/h。
12 h至开始诱导:温度28℃,转速750 r/min,通气量1 NM3/h。开始诱导至诱导10 d:温度23℃,转速750 r/min,通气量1 NM3/h。
1.1.4 主要仪器高速低温离心机和紫外分光光度计(NANO Drop 2000)购自上海赛默飞世尔科技公司。全温震荡培养箱(211B)购自上海智城分析仪器制造有限公司。细胞电转化仪购自Invitrogen公司。10 L微生物全自动发酵罐(Bio-10JS)和甲醇检测流加控制器(FC2002)购自上海保兴生物设备有限公司。XK26层析柱、Typhoon FLA 7000 IP激光扫描成像仪和ImageQuant LAS 4000mini超灵敏化学发光成像仪购自美国GE公司。HD-3000型紫外检测仪购自上海嘉鹏科技有限公司。ELx800酶标仪购自美国Biotek reader公司。
1.2 方法 1.2.1 HSA-ONC融合基因的序列优化与载体构建通过GenBank查询HSA(NM_000477.5)基因序列和ONC基因序列(AF332139.1),选取两者的的成熟肽碱基序列作为融合基因序列,于HSA和ONC(按何庆等[12]方法突变糖基化位点)间插入(Gly4Ser1)3作为连接肽,对融合蛋白核苷酸序列进行密码子偏好性优化设计并提高GC含量,采用HSA自身信号肽作为融合基因的分泌信号肽,序列全长2 187 bp,优化序列经全基因合成后分别构建至分泌型毕赤酵母表达载体pPIC9、pPIC9K和pPICZα-A,合成及构建由上海生工生物工程有限公司进行。
1.2.2 工程酵母菌的制备与筛选提取重组质粒pPIC9/HSA-ONC、pPIC9K/HSA-ONC和pPICZα-A/HSA-ONC,Pme I分别线性化后电转至新鲜制备的酵母感受态细胞,将pPIC9/HSA-ONC、pPIC9K/HSA-ONC、pPICZα-A/HSA-ONC分别转至X-33、GS115和SMD1168三种感受态中,转化方法参考Invitrogen公司毕赤酵母表达手册进行。其中含抗性载体为pPIC9K(G418)和pPICZα-A(Zeocin),组氨酸缺陷型宿主菌为GS115和SMD1168,载体与宿主菌株组合后需要筛选标记方可进行阳性克隆筛选,实际得到以下7种载体-宿主菌组合,依次为:pPICZα-A/X-33、pPIC9/GS115、pPIC9K/GS115、pPICZα-A/GS115、pPIC9/SMD1168、pPIC9K/SMD1168、pPICZα-A /SMD1168。转化的感受态细胞涂布不同抗性平板进行阳性克隆筛选,其中转化pPIC9、pPIC9K的菌液涂布组氨酸缺陷型平板,转化pPICZα-A菌液涂布于pH7.5、含Zeocin(100 μg/mL)的YPD平板。28℃倒置培养3 d,挑取单克隆进行扩大培养,提取酵母基因组作为模板,使用AOX通用引物进行PCR扩增,产物进行琼脂糖电泳鉴定并测序(苏州金唯智生物科技有限公司)。
随机挑选7个组合中鉴定正确菌种分别接种到10 mL YPG/50 mL三角瓶中,于28℃、250 r/min振荡培养18-24 h(一级活化),当OD600值达到10时,按1%接种至100 mL BMGY /1 L三角瓶中,继续于28℃、280 r/min培养24 h后进行甲醇诱导。每12 h补加一次甲醇(发酵液体积的1%),培养条件调整为23℃、200 r/min。诱导10 d后,发酵液于12 000 r/min离心10 min后保留上清,Bradford法测定上清蛋白浓度,取50 μL上清进行15% SDS-PAGE检测(下同),用Typhoon FLA 7000 IP激光扫描成像仪对SDS-PAGE电泳中蛋白条带进行灰度扫描,依据目的条带所占比例进行定量,确定表达量最高的载体-宿主菌组合。
1.2.3 摇瓶发酵的最优条件试验取表达量最高的载体-宿主菌组合菌株进行一级活化,按1%接种至100 mL BMGY/1L三角瓶中。培养基的pH分别为5.5、6.0、6.5、7.0、7.5。在28℃、300 r/min培养24 h后,加入终浓度为1%(V/V)的甲醇开始诱导。转速调整为200 r/min,诱导温度分别设置为20、23、26℃,每12 h补加一次甲醇,补加体积为发酵液体积的1%。诱导10 d后测定发酵液上清蛋白浓度,取50 μL上清进行SDS-PAGE检测,蛋白条带进行灰度扫描和定量,实验重复3次。
1.2.4 10 L发酵规模下最适培养基试验取上述表达量最高的菌种进行一级活化,取400 mL菌液(OD600值达到20左右)分别加入含有4 L BSM培养基和4 L LBSM培养基的10 L发酵罐中,培养基中分别添加4 mL PTM1,设定发酵条件(依据摇瓶最佳条件),发酵过程中控制pH为7.0(流加氨水和磷酸),调节通气量控制溶氧,必要时通入富氧空气,发酵控制按发酵培养条件进行,当溶氧上升时(培养基中甘油耗尽)按DO关联进行甘油补加,当OD600值达到600左右时停止补甘油。饥饿1 h后开始诱导,每升甲醇含20 mL PTM2,诱导过程中控制DO为20%,温度23℃,甲醇浓度0.25%,诱导10 d后取10 μL发酵上清进行SDS-PAGE,进行灰度扫描和定量,检测rHSA-ONC表达情况。
1.2.5 rHSA-ONC的分离纯化以下步骤非特殊要求,均在4℃或冰上进行,优化的纯化条件如下:双水相萃取:取10 L罐诱导10 d发酵液,于4℃,4 000×g离心30 min收集上清,每升上清加入385 g K2HPO4,搅拌至K2HPO4完全溶解后,边搅拌边缓慢加入372 mL无水乙醇,混匀完全后于分液漏斗中-20℃静置过夜,形成上、中、下三层。上层经0.22 μm滤膜过滤,30 kD膜包超滤除盐、除乙醇后进行DEAE弱阴离子交换层析。
离子交换层析:层析柱型号XK26,层析填料DEAE Sepharose Fast Flow,树脂装填量为150 mL,取100 mg浓缩蛋白上样(约50 mL),以1 mL/min的流速上样至平衡(25 mmol/L Na2HPO4溶液,pH 7.0)后的层析柱(2.5 cm×20 cm),用含25 mmol/L Na2HPO4的不同浓度NaCl溶液进行分段洗脱(pH7.0),紫外检测仪监控全程,SDS-PAGE分析各洗脱段组分,灰度扫描后定量。
1.2.6 Western blotting分析取纯化后rHSA-ONC进行SDS-PAGE,用电转仪将目的蛋白从凝胶转移至PVDF膜上,转膜结束后用脱脂奶粉封闭过夜,分别用HSA山羊多抗一抗与ONC兔抗人一抗于室温孵育1 h,TBST洗涤3次,再分别与对应的辣根过氧化物酶(HRP)标记的HSA兔抗山羊二抗和ONC山羊抗兔二抗室温孵育1 h,TBST洗涤3次,超敏ECL底物发光显色,用ImageQuant LAS 4000mini扫描仪和图像定量软件进行定量分析。
1.2.7 SRB法测定rHSA-ONC对癌细胞活性的影响人肝癌7402细胞的培养基为RPMI-1640培养基,HeLa和RH-35细胞的培养基为DMEM培养基,均含10%的胎牛血清、青霉素(100 U/mL)和链霉素(100 U/mL),用0.25%胰蛋白酶消化对数生长期的细胞,将细胞稀释至5×104个/mL,接种于96孔板中,每孔100 μL,于37℃、5% CO2、饱和湿度条件下培养24 h,弃去培养基,分别加入200 μL含不同浓度rONC的培养基,设置无药物处理的对照组和仅含培养基的调零组。于药物处理细胞24h后SRB法测定细胞活性,每个实验组设置3个复孔,实验重复3次。
2 结果 2.1 重组表达质粒的鉴定对pPIC9/HSA-ONC、pPIC9K/HSA-ONC、pPICZα-A/HSA-ONC重组质粒进行酶切、PCR和测序,如图 1所示。pPIC9/HSA-ONC、pPIC9K/HSA-ONC经PmeⅠ单酶切后出现一个12 000 bp左右条带,pPICZα-A/HSA-ONC经PmeⅠ单酶切后出现一个5 000 bp左右条带。pPIC9/HSA-ONC、pPIC9K/HSA-ONC经Bst B I和Eco R I双酶切后出现一个9 000 bp和一个2 187 bp左右条带,pPICZα-A/HSA-ONC经Bam HI和Eco RI双酶切后出现一个3 000 bp和一个2 187 bp左右条带。利用载体通用引物进行PCR扩增,产物理论大小为2 500 bp,结果与理论一致,测序结果显示,序列与预期基因序列一致。
 |
| 1-4:分别为重组质粒pPIC9/ HSA-ONC、Pme I 单酶切、Bst BI 和Eco RI 双酶切、 PCR 扩增结果;5-8 :分别为重组质粒pPIC9K/ HSA-ONC、Pme I 单酶切、 Bst BI 和Eco RI 双酶切、PCR 扩增结果;9-12 :分别为重组质粒pPICZα-A/ HSA-ONC、 Pme I 单酶切、Bam HI 和Eco RI 双酶切、PCR 扩增;M :DNA maker DL10000 图 1 重组表达质粒鉴定 |
将Pme I线性化后的pPIC9/HSA-ONC、pPIC9K/HSA-ONC、pPICZα-A/HSA-ONC重组质粒分别电转入X-33、GS115、SMD1168感受态细胞,经过涂板-培养-筛选等过程获得单克隆。扩大培养后提取酵母基因组DNA,分别以基因组DNA进行PCR,通过琼脂糖凝胶电泳验证PCR产物,结果如图 2所示。重组菌株的基因组均能扩增出2 500 bp的特异条带和2 200 bp的酵母自身条带(图 2-A)。特异条带的大小与预期一致,测序结果正确,表明HSA-ONC已成功整合入毕赤酵母基因组中。每个组合随机挑取若干阳性克隆进行蛋白表达,取诱导10 d发酵液上清进行SDS-PAGE检测(图 2-B)发现,有蛋白条带高表达,其中位于70与100 kD间蛋白条带分子量与预期相符(HSA+非糖基化ONC的理论分子量为78.3 kD),为目的条带。
 |
| 1:protein ladder;2:pPIC9/GS115;3:pPIC9K/GS115;4:PPICZα-A/GS115;5: PPICZα-A/X33 ;6 :pPIC9/SMD1168 ;7 :pPIC9/SMD1168 ;8 :PPICZα- A/SMD1168 图 2 rHSA-ONC 菌株基因组PCR 电泳图(A)和不同载 体- 菌株组合诱导产物SDS-PAGE(B)鉴定图 |
选取表达量最高组合(pPICZα-A /X-33/HSA-ONC)的菌株在1 L摇瓶规模进行不同培养基pH和诱导温度的表达条件优化,发酵结果如图 3所示,诱导10 d时,rHSA-ONC的最佳表达条件为:23℃、pH7.0,表达量最高为235 mg/L
 |
| 图 3 不同pH 和诱导温度对rHSA-ONC 表达量的影响 |
取表达量最高组合菌株进行扩大培养,当菌体OD600达到20时,取400 mL菌液,接种到分别含4 L BSM和4 L LBSM的10 L发酵罐,培养基pH和温度依据摇瓶最佳结果进行设定。待菌体OD600达到600左右开始补甲醇,诱导10 d后,取10 μL发酵液上清进行SDS-PAGE检测。结果(图 4)表明,在LBSM培养基中诱导10 d时rHSA-ONC表达量最高,达到2.02 g/L。
 |
| 图 4 不同培养基对rHSA-ONC 表达量的影响 |
取10 L罐诱导10 d发酵液,离心收集上清,上清经双水相萃取后形成上、中、下三层,用SDS-PAGE分别检测三层的蛋白分布情况(图 5-B),取rHSA-ONC所在层进行0.22 μm过滤和30 kD超滤。将浓缩样品用DEAE离子交换层析进一步分离纯化,如图 5-A所示,在凝胶层析过程出现3个蛋白洗脱峰(P1、P2和P3)。rHSA-ONC主要分布在洗脱液峰P2中,灰度扫描后进行定量分析,rHSA-ONC的纯度大于95%,收率高于70%(图 5-B)。纯化后样品经Western blot检测发现,rHSA-ONC呈现HSA和ONC双阳性(图 5-C)。
 |
| (A)P1 :平衡液洗脱;P2 :25 mmol/L Na2HPO4+75mNaCl 洗脱;P3 :25 mmol/L Na2HPO4+1M NaCl 洗脱。(B)1:10 μg BSA;2:上清;3:双水相上相; 4 :双水相中相;5 :双水相下相;6 :超滤浓缩液。(C)M :蛋白maker ;1 : HSA 抗体检测rHSA-ONC ;2 :ONC 抗体检测rHSA-ONC 图 5 rHSA-ONC 离子交换层析检测谱图(A)、SDSPAGE 纯度鉴定(B)及Western blotting 鉴定(C) |
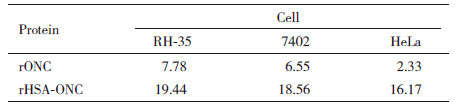
不同浓度的rHSA-ONC处理细胞后,SRB法检测细胞的活力,结果(图 6)表明,6-24 μmol/L的rHSA-ONC处理RH-35、7402和HeLa细胞后24 h,细胞增殖明显受到抑制,表明rHSA-ONC能抑制多种肿瘤细胞增殖且呈剂量依赖效应,对HeLa、7402、RH-35细胞的IC50依次为:19.44、18.56、16.17 μmol/L,与非糖基化rONC IC50相比,活性发生明显下降(表 1)。
 |
| 图 6 rHSA-ONC 对RH-35、7402、HeLa 细胞的抑制作 用图 |
宿主系统与外源基因密码子的差异性经常造成基因沉默或蛋白表达量很低,对外源基因进行密码子优化以适应宿主系统将有利于外源蛋白的表达[17, 18, 19],对酵母偏好性密码子的应用及发酵过程中控制条件的优化是近几年提高外源蛋白表达量和活性的常见手段,它们的应用也为外源蛋白在酵母系统中的表达带来了新的发展[20, 21]。
将两个不同的基因片段通过一段合适的连接肽进行连接,可直接表达融合蛋白,融合蛋白通常具有双重活性,但融合蛋白较原有蛋白会在功能与活性上发生改变[22, 23]。融合蛋白间引入的连接肽过长则可能增加蛋白的免疫原性,还可能影响目的蛋白的活性和功能;而过短的链间连接肽会在分子间造成空间效应,可能影响蛋白的正确折叠[24, 25];因此应当选择能够有效分隔融合蛋白两个功能基团的最短连接肽。一般来说,3-5个氨基酸的连接肽可满足大多数融合蛋白的正确折叠需求,长度不宜超过15个氨基酸[26, 27, 28],连接肽的柔性和疏水性能保证蛋白质的功能结构域不受影响[29]。
为获得高表达的非糖基化rONC,本文将ONC(突变掉糖基化位点)与HSA的C端连接后进行表达,采用目前普遍使用的柔性连接肽,即(Gly4Ser1)3作为融合蛋白间的连接肽,简称HSA-ONC。首先利用酵母密码子偏好性对融合序列进行优化,同时提高序列GC含量,采用HSA的自身信号肽作为分泌信号肽,构建3种表达载体pPIC9/HSA-ONC、pPIC9K/HSA-ONC、pPICZα-A/HSA-ONC,分别转化酵母菌X-33、GS115和SMD1168,筛选得到7种载体-受体菌组合,初步诱导后发现表达量最高的载体-受体菌组合为pPICZα-A/X-33/HSA-ONC,在摇瓶规模的最佳表达条件(23℃、pH 7.0)下,诱导10 d时rHSA-ONC的表达量达到235 mg/L。在10 L规模下使用LS-BSM培养基诱导10 d时,rHSA-ONC的最高表达量为2.02 g/L,是等摩尔量条件下非糖基化rONC表达量[12]的2倍,但仍低于rHSA的表达量(10 g/L)。经多次优化纯化条件后,采用双水相偶联DEAE离子交换层析方法进行纯化,最终得到了收率高于70%、纯度≥95%的rHSA-ONC。MTT法检测发现rHSA-ONC能杀伤多种癌细胞,与文献报道结果[9, 10, 11, 12]的ONC作用一致,但对肿瘤细胞的杀伤作用较rONC下降60%。
本实验优化表达条件后获得了较高表达量的rHSA-ONC(2.02 g/L),但仍明显低于rHSA的表达量,经电泳检测发现,蛋白积累过程中有较多的目的蛋白发生降解,目的蛋白的积累减少与之有关,若能增强融合蛋白的稳定性,减少其降解,目的蛋白的表达量可至少提高1倍。rHSA-ONC的癌细胞杀伤作用与rONC相比下降明显,可能与HSA分子量较大,空间上干扰了rONC的折叠,进而影响其活性。由此可见,蛋白的稳定性与活性与融合蛋白间连接肽密切相关,我们将在本实验基础上进一步研究不同长度或不同类型连接肽对融合蛋白的影响,以得到表达量、稳定性明显提高,生物学活性较强的rHSA-ONC,为降低ONC的生产成本及产业化奠定基础。
4 结论本研究首次将HSA与ONC(非糖基化)融合后在真核系统进行重组表达,利用HSA的表达优势和不同载体-宿主组合筛选得到高表达组合,优化表达条件后最终表达量达到2.02 g/L,是等摩尔量条件下非糖基化rONC表达量的2倍,通过双水相偶联柱层析快速高效地进行纯化。运用SRB法测定rHSA-ONC对癌细胞的抑制活性,rHSA-ONC能杀伤多种癌细胞,但活性低于rONC。
| [1] | Porta C, Paglino C, Mutti L. Ranpirnase and its potential for the treatment of unresectable malignant mesothelioma[J]. Biologics, 2008, 2(4):601-609. |
| [2] | Darzynkiewicz Z, Carter SP, Mikulski SM, et al. Cytostatic and cytotoxic effects of Pannon(P-30 Protein), a novel anticancer agent[J]. Cell Tissue Kinet, 1988, 21(3):169-182. |
| [3] | Ardelt W, Mikulski SM, Shogen K, et al. Amino acid sequence of an anti-tumor protein from Rana pipiens oocytes and early embryos. Homology to pancreatic ribonucleases[J]. J Biol Chem, 1991, 266(1):245-251. |
| [4] | 田雪, 王庆诚, 沈如凌, 等. Onconase研究与开发的最新进展[J]. 中国细胞生物学报, 2010, 32(6):927-934. |
| [5] | Leland PA, Schultz LW, Kim BM, Raines RT. Ribonuclease A variants with potent cytotoxic activity[J]. Proc Nat Acad Sci USA, 1998, 95(18):10407-10412. |
| [6] | Iohnson RJ, Chao TY, Lavis LD, et al. Cytotoxic ribonucleases:The dichotomy of coulombic forces[J]. Biochemistry, 2007, 46(36):10308-10316. |
| [7] | Chang CH, Gupta P, Michel R, et al. Ranpirnase(frog RNase)targeted with a humanized, internalizing, anti-Trop-2 antibody has potent cytotoxicity against diverse epithelial cancer cells[J]. Mol Cancer Ther, 2010, 9(8):2276-2286. |
| [8] | Qiao M, Zu LD, He XH, et al. Onconase downregulates microRNA expression through targeting microRNA precursors[J]. Cell Res, 2012, 22(7):1199-1202. |
| [9] | 汪楠, 唐小军, 熊四平, 等. 重组豹蛙酶的制备及其生物学特性分析[J]. 南京医科大学学报, 2013, 33(8):1034-1038. |
| [10] | 胡晓珺. 核糖核酸酶Onconase及其融合蛋白表达[D]. 上海:华东理工大学, 2012. |
| [11] | 郭红, 徐殿胜, 王庆诚. 核糖核酸酶Onconase的稀释复性条件优化研究[J]. 化学与生物工程, 2013, 10(30):57-60. |
| [12] | 何庆. 糖基化和非糖基化Onconase在巴斯德毕赤酵母中的分泌表达、纯化与活性鉴定[D]. 北京:中国人民解放军军事医学科学院, 2009. |
| [13] | 贾茜, 刘文献, 赵伟, 等. 一种重组人血清白蛋白的纯化方法及其应用:中国, 101768206A, 1[P]. 2008-12-30. |
| [14] | 徐存拴, 史世领, 史世会, 等. 一种表达人血清白蛋白的质粒和重组菌以及它们的应用:中国, 103194482A, 1[P]. 2013-3-12. |
| [15] | Kratz F. Albumin as a drug carrier:design of prodrugs, drug conj-ugates and nanoparticles[J]. J Control Release, 2008, 132(3):171-183. |
| [16] | 王康, 王强, 张磊. SRB 法和MTT法检测PDGF-BB对牛眼小梁细胞增殖的影响[J]. 滨州医学院学报, 2012, 35(1):10-12. |
| [17] | Mellitzer A, Weis R, Glieder A, et al. Expression of lignocellulolytic enzymes in Pichia pastoris[J]. Microb Cell Fact, 2012, 2012(11):61-72. |
| [18] | Sygmund C, Gutmann A, Krondorfer I, et al. Simple and efficient expression of Agaricus meleagris pyranose dehydrogenase in Pichia pastoris[J]. Appl Microbiol Biotechnol, 2012, 94(3):695-704. |
| [19] | Abad S, Nahalka J, Winkler M, et al. High-level expression of Rhodotorula gracilis D-amion acid oxidase in Pichia pastoris[J]. Biotechnol Lett, 2011, 33(3):557-563. |
| [20] | Spatz SJ, Volkening JD, Mullis R, et al. Expression of chicken parvovirus VP2 in chicken embryo fibroblasts require codon optimization for production of naked DNA and Vecored meleagrid herpesvirus type 1 vaccine[J]. Virus Genes, 2013, 47(2):259-267. |
| [21] | Park S, Pack SP, Lee J. Expression of codon-optmiazed phosphoen- olpyruvate carboxylase gene from Glaciecola sp. HTCC2999 in Es-cherichia coli and its application for C4 chemical production[J]. Appl Biochem Biotech, 2012, 167(7):1845-1853. |
| [22] | Geng SS, Feng J, Li Y, et a1. Binding activity difference of anti-CD20 scFv-Fc fusion protein derived from variable domain exchange[J]. Cell Mol Immunol, 2006, 3(6):439-443. |
| [23] | Tranchant I, Herve AC, Carlisle S, et a1. Design and synthesis of ferrocene probe molecules for detection by electrochemical methods[J]. Bioconjug Chem, 2006, 17(5):1256-1264. |
| [24] | Gustavsson M, Lehtiö J, Denman S, et a1. Stable linker peptides for a cellulose-binding domain-lipase fusion protein expressed in Pichia pastoris[J]. Protein Eng, 2001, 14(9):711-715. |
| [25] | Le Gall F, Reusch U, Little M, et a1. Effect of linker sequences between the antibody variable domains on the formation, stability and biological activity of a bispecific tandem diabody[J]. Protein Eng Des Sel, 2004, 17(4):357-366. |
| [26] | Hu W, Li F, Yang X, et al. A flexible peptide linker enhances the immunoreactivity of two copies HBsAg preSl(21-47)fusion protein[J]. J Biotechnol, 2004, 107(1):83-90. |
| [27] | Ha SH, Park JJ, Kim JW, et al. Molecular cloning and high-level expression of G2 protein of hantaan(HTN)virus 76-118 strain in the yeast Pichia pastoris KM71[J]. Virus Genes, 2001, 22(2):167-173. |
| [28] | Arai R, Ueda H, Kitayama A, et al. Design of the linkers which effectively separate domains of a bifunctional fusion protein[J]. Protein Eng, 2001, 14(8):529-532. |
| [29] | Wriggers W, Chakravarty S, Jennings PA. Control of protein functional dynamics by peptide linkers[J]. Biopolymers, 2005, 80(6):736-746. |