



b. Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla 666303, China;
c. University of Chinese Academy of Sciences, Beijing 100049, China;
d. State Key Laboratory of Paleobiology and Stratigraphy, Nanjing Institute of Geology and Paleontology, Chinese Academy of Sciences, Nanjing 210008, China
Eurya Thunb. is a large genus of the tea family Theaceae, comprising approximately 130 species (Min and Bartholomew, 2007). Its extant species are mainly distributed in tropical to subtropical Asia, and in the western and southern Pacific Islands (Min and Bartholomew, 2007), with a few species in tropical regions of the New World (Łańcucka-Środoniowa, 1966). They are evergreen shrubs, small trees, and rarely large trees, commonly found as dominant understory elements in tropical to subtropical broadleaved evergreen forests (WGVY, 1987; Min and Bartholomew, 2007).
Although Eurya is not distributed in Europe now, its fossil record is especially rich in this continent, where at least 12 extinct species from the Late Cretaceous to early Pleistocene have been documented (Mai, 1960, 1971; Knobloch, 1977; Łańcucka-Środoniowa, 1981; Friis, 1985; Knobloch and Mai, 1986; van der Burgh, 1987; Friis et al., 2011; Martinetto et al., 2015). Unlike Europe, the fossil occurrences of Eurya in East Asia are rather scarce (Tanai and Uemura, 1991; Momohara, 1992; Momohara and Saito, 2001; Yamakawa et al., 2015), even though this region shows the highest species richness of the genus today. Although molecular data can explain the emergence and diversification of Eurya (Wu et al., 2007), the lack of fossil evidence limits further understanding of the evolution and establishment of its high modern species diversity in East Asia.
In this study, we report a new fossil record of Eurya based on three-dimensionally preserved seeds from the late Pliocene of northwestern Yunnan, southwestern China. We perform detailed morphological and anatomical examinations of these fossil seeds, and discuss the paleobiogeographic implications of this new fossil finding.
2 Materials and methods 2.1 Fossil site and geological horizonThe present fossil seeds were collected from deposits at Nanbanbang Village, Heqing Basin, northwestern Yunnan Province (26°310 N, 100°100 E; 2200 m a.s.l.; Fig. 1). The Heqing Basin is located at the southeastern fringe of the Qinghai-Tibet Plateau within the renowned Hengduan Mountains. It is a tectonic default basin surrounded by mountains with a distinguishable vertical vegetation belt spectrum (Shen et al., 2007). The local climate is primarily influenced by the Indian Monsoon, which is characterized by warm, wet summers and cool, dry winters (Xiao et al., 2010; An et al., 2011). The section studied is unconformably overlain by Quaternary deposits and is composed of horizontally laminated carbonaceous layers imbedded by fine gray sandstones. These carbonaceous layers contain abundant plant remains, mainly fruits and seeds. The age of the carbonaceous layers was previously assigned to the early Pleistocene (Zhu et al., 2016). The latest geological survey shows that the underlying sedimentary layers beneath these carbonaceous layers contain abundant leaf fossils dominated by evergreen sclerophyllous oaks (Quercus sect. Heterobalanus). In northwestern Yunnan, the dominance of evergreen sclerophyllous oaks in a fossil flora is a common indicator of the Sanying Formation (Tao, 1986), which has been determined to be the late Pliocene based on stratigraphic correlations, palynological information, mammal fossils, and paleomagnetic data (Tao and Kong, 1973; Tao, 1986; Ge and Li, 1999; Su et al., 2011; Li et al., 2013). Therefore, the age of the present fossil seeds and fruits is assigned to the late Pliocene.

|
| Fig. 1 Map showing the fossil site (a, b) and outcrops (c) from which the present fossil seeds were collected. |
More than 1000 specimens of charcoalified fruits and seeds were collected from the fossil site. Among them, more than 80 seeds and seed fragments of Eurya were identified through observations under a binocular microscope (Leica, S8AP0). The fossil seeds were cleaned by an ultrasonic cleaner at 40 kHz (KO-50M) for 5-10 s. Air dried, they were then observed under a 3D Super Depth Digital Microscope (ZEISS Smartzoom 5) and images were taken. Five seed specimens were further studied under a scanning electron microscope (SEM, Zeiss EVO LS10) both morphologically and anatomically. For comparative analysis, extant seeds of Eurya obtained from herbarium specimens housed at the Herbarium of Kunming Institute of Botany (KUN) were also examined using the same procedure as the fossils. The descriptive terminology mainly follows Friis (1985). All studied fossil specimens are numbered and kept at the KUN.
3 SystematicsFamily: Theaceae Mirb., 1816
Genus: Eurya Thunb., 1783
Species: Eurya stigmosa (Ludwig) Mai, 1960
Fossil specimens: NBB 020 (Plate Ⅰ, 1), NBB 021 (Plate Ⅰ, 2), NBB 022 (Plate Ⅰ, 3), NBB 023 (Plate Ⅰ, 4), NBB 024 (Plate Ⅰ, 5), NBB 025 (Plate Ⅰ, 6), NBB 026 (Plate Ⅰ, 7), NBB 027 (Plate Ⅰ, 8), NBB 028 (Plate Ⅰ, 9), NBB 029 (Plate Ⅰ, 10), NBB 030 (Plate Ⅰ, 11), NBB 031 (Plate Ⅰ, 12), NBB 032 (Plate Ⅱ, 1, 8), NBB 033 (Plate Ⅱ, 2), NBB 034 (Plate Ⅱ, 3), NBB 035 (Plate Ⅱ, 4, 6, 7), NBB 036 (Plate Ⅱ, 5, 9, 10, 11, 12), and NBB 037-072.

|
| Plate Ⅰ. Fossil seeds of Eurya stigmosa from Nanbanbang under the 3D Super Depth Digital Microscope. Scale bars=0.5 mm for all images. T=tegmen. |

|
| Plate Ⅱ. Fossil seeds of Eurya stigmosa from Nanbanbang under the SEM. Scale bars=0.25 mm for 6 and 8; 0.05 mm for 7, 9, and 12; 0.1 mm for 5 and 11; and 0.5 mm for 1-4, and 10. 1-4. General shape of the fossil seeds. 5. Longitudinal section of the seed wall showing the two distinct layers consisting of large cells of the exotesta and small cells of the endotesta. 6. External surface of the seed showing the concentrically arranged cells. 7. Details of the cells on the seed external surface, showing the funnel-shaped and strongly thickened cells of the testa with finely pitted walls. 8. Cells on the center part of the seed external surface, showing the compact, less visible cell lumens and thicker walls. 9. Internal surface of the seed showing elongated elliptic and slightly thickened cells, which are concentrically arranged around the condyle. 10. Internal view of the seed showing the reniform or horseshoe-shaped embryo cavity, reversed and slightly curved V-shaped condyle, subtriangular raphe cavity, micropyle, hilum, and chalaza. 11, 12. Details of the condyle showing the clavate cells. ex=exotesta, en=endotesta, c=condyle, ch=chalaza, h=hilum, m=micropyle, r=raphe. |
Fossil locality: Nanbanbang Village, Heqing County, northwest Yunnan Province, southwest China (26°310 N, 100°100 E; 2200 m a.s.l.)
Stratigraphic horizon: The upper Pliocene Sanying Formation Description: Seeds are slightly campylotropous, laterally flattened, and subcircular to slightly angular in outline (Plate Ⅰ). They are 1.07-2.02 mm in length and 1.03-1.96 mm in width, with a lengthewidth ratio of 0.82-1.32. The seed surface is foveolate with funnel-shaped cells (Plate Ⅱ, 1-4, 6), which are concentrically arranged around the condyle-raphe region in 8-12 rows parallel to the seed margin (Plate Ⅱ, 1-4, 6-8). The cells are 0.03-0.15 mm (0.08 mm on average) in diameter, with thickened and finely pitted inner periclinal and anticlinal walls (Plate Ⅱ, 7), and with cell lumens that gradually narrow towards the base with polygonal facets (Plate Ⅱ, 6-7). The cavity of each seed valve is subdivided by a characteristic reversed and slightly curved Vshaped condyle into a reniform or horseshoe-shaped embryo cavity and raphe (Plate Ⅱ, 10). The condyle consists of two limbs with equal length but unequal width, which extend from the seed base to almost half of the seed width (Plate Ⅱ, 10). The raphe is marked by a subtriangular cavity, and is formed by the encompassment of the condyle and seed basal margin. The hilar scar is oblong and located between the raphe and micropyle. The seed internal surface is covered by elongated elliptic and slightly thickened cells, which are concentrically arranged around the condyle in rows parallel to the seed margin (Plate Ⅱ, 9-10). The seed wall is 0.06-0.16 mm thick, sclerotic, composed of a thick exotesta, with large cells, and a thin endotesta, with small cells (Plate Ⅱ, 5). The thin, membranous remains of the tegmen can be observed attached to the external surface of testa in several seeds (Plate Ⅰ, 12).
Morphological comparisons: Although seeds of Eurya are seemingly similar to those of its relatives, such as Adinandra W. Jack, Cleyera Thunberg, and Freziera Willd, they can be reliably distinguished from these genera by key structural details (Friis, 1985; Table 1). Generally, Eurya seeds are characterized by a reticulum of funnel-shaped cells with polygonal facets on the external seed surface (Friis, 1985). This diagnostic feature is consistent with our fossil seeds, which differs from the other three genera. Moreover, our fossils also share some other important anatomical features with extant Eurya seeds, such as the reniform or horseshoe-shaped embryo cavity, the prominent condyle in the seed valve, and the position of raphe and hilum (Friis, 1985). All of these traits suggest that our fossil seeds can be ascribed to Eurya. Among the examined modern seeds of Eurya, Eurya yunnanensis, a species presently found mainly in western and southeastern Yunnan, shows the greatest resemblance to the fossil seeds under study. However, the fossil seeds are distinct from this species by their slightly larger size, finely pitted cells on the seed external surface, and the difference in cell types on the condyle and seed internal surface (Plates Ⅱ and Ⅲ).
| Genus | Seed general shape | Exotesta | Endotesta | Seed draft | ||
| Cell shape | Cell wall | Layers of crystal cells | Cell wall | |||
| Adinandra | Slightly campylotropous | Not funnel-shaped | Equally thick | One to several layers | Often thick | 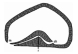 |
| Cleyera | Strongly campylotropous, usually larger than those of the other three genera | Not funnel-shaped | Equally thick | Usually one layer | - |  |
| Eurya | Slightly campylotropous, or more rarely anatropous | Funnel-shaped | Strongly thickened | One or two layers | Rather thin |  |
| Freziera | Usually anatropous or slightly campylotropous | Not funnel-shaped | Equally thick | Several layers | Strongly thickened |  |

|
| Plate Ⅲ. Extant seeds of Eurya yunnanensis under the SEM. Scale bars=0.25 mm for 3 and 5; 0.05 mm for 1, 4, 7, and 8; and 0.5 mm for 2 and 6. 1. Longitudinal section of the seed wall, showing the two distinct layers consisting of large cells of the exotesta and small cells of the endotesta. 2. General shape of the seed and the cells concentrically arranged on the external surface. 3, 4. Details of the cells on the seed external surface, showing the funnel-shaped cells of the testa with smooth surface. 5. The condyle structure and the elongated polygonal cells on the internal surface of the seed which are concentrically arranged around the condyle. 6. Internal view of the seed showing the reniform or horseshoeshaped embryo cavity, reversed and slightly curved V-shaped condyle, subtriangular raphe cavity, micropyle, hilum, and chalaza. 7. Details of the condyle showing the elliptic or subcircular cells. 8. Details of the cells on the internal surface of the seed, showing the thin and raised cell wall. ex=exotesta, en=endotesta, c=condyle, ch=chalaza, h=hilum, m=micropyle, r=raphe. |
Fossil seeds of Eurya that have previously been described are largely from Europe (Chandler, 1963; Mai, 1971; Knobloch, 1977; Łańcucka-Środoniowa, 1981; Friis, 1985; Knobloch and Mai, 1986; van der Burgh, 1987; Martinetto et al., 2015), and to a limited extent from Japan in East Asia (Momohara, 1992; Momohara and Saito, 2001; Yamakawa et al., 2015) and Nepal in South Asia (Bhandari et al., 2009, 2010). They are assigned to 12 fossil species and one modern species (Table 3). We compared our fossil seeds with these 12 fossil species using four key characteristics: size of the cells on the seed external surface, seed size, seed shape, and testa thickness (Table 2). Our fossil seeds most resemble the seeds of E. stigmosa (Ludwig) Mai reported from Paleocene to early Pleistocene of Europe (Table 3), that have similarly large cells on the seed external surface. Our fossils also have similar seed size (1.07-2.02×1.03-1.96 mm) and testa thickness (0.06-0.16 mm) with those of E. stigmosa. Other important seed characteristics, such as the subcircular to slightly angular seed shape, are also shared by our fossils and this species. We have therefore treated our fossils as belonging to E. stigmosa.
| Taxa | Cells on the external surface | Seed size | Shape | Testa | References |
| E. stigmosa (Ludwig) Mai | Large, 0.03-0.15 (0.08 on average) mm in diameter | Medium, 1.07-2.02 × 1.03-1.96(1.47 × 1.45 on average) mm | Subcircular, campylotropous, laterally flattened | Thick, 0.06-0.16 mm | This study |
| E. mudensis Chandler | Large, 0.05-0.2(0.1onaverage)mm in diameter | Small, 1.0-1.5 mm in diameter | More rounded | Thick, brittle | Mai (1971) |
| E. stigmosa (Ludwig) Mai | Large cavites, 0.05-0.2 (0.1 on average) mm in diameter | Medium, 1.0-2.5 (rarely 3)mm in diameter | Great variable (reniform, broadly oval, sub-triangular, angular to almost circular) | Thick, brittle, 0.05-0.15 mm | Mai (1971) and Friis (1985) |
| E. poolensis (Chandler) Mai | Large, 0.05-0.2 (0.1 on average) mm in diameter | Medium, 1.9-2.3 mm in diameter | Very oblique oval | Thick, brittle | Mai (1971) |
| E. lusatica Mai | Large cavites, 0.05-0.2 (0.1 on average) mm in diameter | Large, 2.0-3.2 mm in diameter | Subcircular to obliquely oval | Thick, brittle | Mai (1971) |
| E. becktonensis Chandler | Large, 0.05-0.2 (0.1 on average) mm in diameter | Large, 1.8-4.2 mm in diameter | Elongated oval to obliquely triangular, somewhat angular | Thick, brittle | Mai (1971) |
| E. maii Knobloch | Large, 0.06-0.13 mm in diameter, eccentric, large (sometimes heptagonal) polygons | Medium, 1.22-2.02 × 0.97-2.0 (1.62 × 1.38 on average) mm | Roundish-lenticular to oval, more or less compressed | Excessively thick, 0.13-0.22 mm | Knobloch (1977) |
| E. boveyana (Chandler) Mai | Fine, 0.02-0.08 (0.05 on average) mm in diameter | Medium, 1.0-2.0 mm in diameter | Subcircular to oval, more or less rounded, always compressed | Thin, elastic | Mai (1971) |
| E. obliqua (Chandler) Mai | Fine, 0.02-0.08 (0.05 on average) mm in diameter | Small, 1.25-1.7 mm in diameter | Rounded triangular to irregular, always somewhat angular | Thick, brittle, strongly thickened in the central area | Mai (1971) |
| E. dubia (Chandler) Mai | Fine, 0.02-0.08 (0.05 on average) mm in diameter | Medium, 1.25-2.0 mm in diameter | Rounded triangular to irregular, always somewhat angular | Thick, brittle, strongly thickened in the central area | Mai (1971) |
| E. lentiformis (Chandler) | Mai Fine, 0.02-0.08 (0.05 on average) mm in diameter | Large, about 2.5 mm in diameter | R ounded triangular to irregular, always somewhat angular | Thick, brittle, strongly thickened in the central area | Mai (1971) |
| E. crassitesta Knobloch | Fine, about 0.05 mm in diameter, concentric, equal, rounded and annular thickened polygons | Small, 0.8-1.4 × 0.7-1.8 mm | Rounded | Thick, 0.08-0.8 mm | Knobloch (1977) |
| E. holyi Knobloch | Fine, eccentric, equally sized polygons | Small, 0.75-1.33 × 0.55-0.97 (1.02 × 0.8 on average) mm | Semicircular, spherical, broadly elliptic, rarely rounded | Thick, 0.08-0.11 mm | Knobloch (1977) |
| Taxa | Age | Fossil type | Locality | References |
| E. sp. | Late Pleistocene | Seed | Central Nepal | Bhandari et al.(2009, 2010) |
| E. stigmosa (Ludwig) Mai | Late Pliocene | Seed | NW Yunnan, SW China | This study |
| E. japonica Thunberg | Late Pliocene | Seed | Central Japan | Yamakawa et al. (2015) |
| E. sp. | Late Pliocene | Seed | SW Japan | Momohara (1992) |
| E. stigmosa (Ludwig) Mai | Early Pliocene-Early Pleistocene | Seed | N Italy | Martinetto et al. (2015) |
| E. stigmosa (Ludwig) Mai | Middle Pliocene | Seed | Central Italy | Martinetto (2001) |
| E. stigmosa (Ludwig) Mai | Middle Pliocene | Seed | NW Italy | Basilici et al. (1997) |
| E. stigmosa (Ludwig) Mai | Early Pliocene | Seed | NW Italy | Martinetto et al. (1997) |
| E. stigmosa (Ludwig) Mai | Pliocene | Seed | Germany | van der Burgh(1978, 1983) |
| E. sp. | Late Miocene to Early Pliocene | Seed | England | Boulter (1971) |
| E. ciliata Merrill | Late Miocene to Early Pliocene | Leaf | SW Yunnan, SW China | Tao and Du (1982) |
| E. stigmosa (Ludwig) Mai | Late Miocene to Early Pliocene | Seed | SW Poland | Dyjor et al. (1992) |
| E. sp. | Late Miocene | Leaf | Japan | Ozaki (1991) |
| E. lusatica Mai | Late Miocene | Seed | Eschweiler, Germany | van der Burgh (1987) |
| E. japonica Thunberg | Late Miocene | Seed | Central Japan | Momohara and Saito (2001) |
| E. stigmosa (Ludwig) Mai | Late Miocene | Seed | Eschweiler, Germany | van der Burgh (1987) |
| E. stigmosa (Ludwig) Mai | Middle Miocene | Seed | Denmark | Friis(1979, 1985) |
| E. stigmosa (Ludwig) Mai | Early Miocene | Seed | Czech | Teodoridis and Kvacek (2006) |
| E. stigmosa (Ludwig) Mai | Early Miocene | Seed | Austria | Meller (1998) and Meller et al. (1999) |
| E. stigmosa (Ludwig) Mai | Miocene | Seed | South Poland | Łańcucka-Środoniowa(1966, 1981, 1984) |
| E. stigmosa (Ludwig) Mai | Miocene | Seed | Germany | Gregor (1982) |
| E. stigmosa (Ludwig) Mai | Miocene | Seed | Germany | Mai (1960) |
| E. lusatica Mai | Miocene | Seed | Puschwitz, Germany | Mai (1971) |
| E. aff. acuminatissima Merr. & Chun | Late Oligocene | Leaf | SE Bulgaria | Bozukov and Palamarev (1995) and Bozukov et al. (2008) |
| E. sp. | Late Oligocene | Leaf | Japan | Tanai and Uemura (1991) |
| E. dubia (Chandler) Mai | Middle Oligocene | Seed | Devonshire, England | Mai (1971) |
| E. dubia (Chandler) Mai | Middle Oligocene | Seed | Nerchau, Germany | Mai (1971) |
| E. mudensis Chandler | Middle Oligocene | Seed | Germany | Mai (1971) |
| E. stigmosa (Ludwig) Mai | Middle Oligocene | Seed | Devonshire, England | Mai (1971) |
| E. boveyana (Chandler) Mai | Middle Oligocene | Seed | Devonshire, England | Mai (1971) |
| E. boveyana (Chandler) Mai | Oligocene | Seed | Germany | Mai and Walther (1978) |
| E. boveyana (Chandler) Mai | Late Eocene to early Oligocene | Seed | Germany | Mai (1971) |
| E. sp. | Late Eocene | Seed | Russia | Nikitin (2006) |
| E. lentiformis (Chandler) Mai | Late Eocene | Seed | Wight, England | Mai (1971) |
| E. becktonensis Chandler | Late Eocene | Seed | Hants, England | Mai (1971) |
| E. mudensis Chandler | Late Eocene | Seed | England | Chandler (1963) and Mai (1971) |
| E. obliqua (Chandler) Mai | Middle Eocene | Seed | Egeln, Germany | Mai (1971) |
| E. poolensis (Chandler) Mai | Middle Eocene | Seed | Bournemouth, England | Mai (1971) |
| E. stigmosa (Ludwig) Mai | Middle Eocene | Seed | Bournemouth, England | Mai (1971) |
| E. obliqua (Chandler) Mai | Early Eocene | Seed | Dorset, England | Mai (1971) |
| E. stigmosa (Ludwig) Mai | Early Eocene | Seed | Oscherslebener Mulde, Germany | Mai (1971) |
| E. stigmosa (Ludwig) Mai | Paleocene | Seed | Reading Beds, England | Mai (1971) |
| E. mudensis Chandler | Paleocene | Seed | Germany | Mai (1971) |
| E. crassitesta Knobloch | Late Cretaceous to Paleocene | Seed | Moravian Carpathian, Czech Republic | Knobloch (1977) |
| E. maii Knobloch | Late Cretaceous to Paleocene | Seed | Moravian Carpathian, Czech Republic | Knobloch (1977) |
| E. holyi Knobloch | Late Cretaceous to Paleocene | Seed | Moravian Carpathian, Czech Republic | Knobloch (1977) |
| E. crassitesta Knobloch | Middle-Late Cretaceous | Seed | Austria | Knobloch and Mai(1986, 1991) |
Eurya has a rich fossil record, represented primarily by seed remains and a few leaf impressions (Tao and Du, 1982; Ozaki, 1991; Tanai and Uemura, 1991; Bozukov et al., 2008). The oldest known records of the genus are seed remains from the middle to Late Cretaceous in Austria (Knobloch and Mai, 1986, 1991) and from the Late Cretaceous to Paleocene in the Czech Republic, central Europe (Knobloch, 1977). This may imply a central European origin. Fossil record also indicates that Europe, where about 12 known fossil species of the genus have been reported, may have been a center for Eurya evolution and diversification in the past (Chandler, 1963; Mai, 1971; Knobloch, 1977; Łańcucka-Środoniowa, 1981; Gregor, 1982; Friis, 1985; Knobloch and Mai, 1986; van der Burgh, 1987; Martinetto et al., 2015) (Table 3; Fig. 2). This hypothesis may be consistent with the warm and humid climate throughout the European Cenozoic (Mosbrugger et al., 2005). Based on the lack of fossil occurrences in the continent after the early Pleistocene, the genus likely disappeared from Europe by that time (Martinetto et al., 2015). This may be largely attributed to the more severe effect of the Quaternary glaciations on Europe compared with relatively low latitude regions (Ehlers et al., 2011).

|
| Fig. 2 Spatial distribution of modern (a) and fossil (b) species of Eurya. |
In contrast to Europe, East Asia has revealed a much lower diversity of Eurya in its geological past. Despite flourishing in the modern era, there are only six documented fossil occurrences of Eurya in East Asia (Tao and Du, 1982; Ozaki, 1991; Tanai and Uemura, 1991; Momohara, 1992; Momohara and Saito, 2001; Yamakawa et al., 2015). The oldest known fossil record is dated to the late Oligocene (Tanai and Uemura, 1991), implying that Eurya probably first appeared in East Asia no later than the late Oligocene. The newly described fossil occurrence represents the first seed fossil record of Eurya in China, and one of the few fossil records of the genus in East Asia. As E. stigmosa has been frequently documented from the Paleocene to early Pleistocene of Europe (Mai, 1971; Łańcucka-Środoniowa, 1981; Gregor, 1982; Friis, 1985; van der Burgh, 1987; Teodoridis and Kvaček, 2006; Martinetto et al., 2015), it may indicate a close affinity between East Asian and European species of the genus. It may suggest the ancient species of East Asian Eurya originated in Europe. The genus may have further dispersed to other regions of Asia, as some other fossil occurrences of Eurya are also reported from the continent, e.g., the late Miocene and late Pliocene of Japan (Ozaki, 1991; Yamakawa et al., 2015), and the late Pleistocene of Nepal (Bhandari et al., 2009, 2010). These ossils may represent partial histories of the ancient lineages of modern Eurya in Asia.
AcknowledgmentsWe thank Lin-Bo Jia from Kunming Institute of Botany, Chinese Academy of Sciences, for help with fossil collection; and the editor and two anonymous reviewers for improving the manuscript. Fossil examinations with the 3D Super Depth Digital Microscope and SEM were performed in the Central Lab of Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences; and examination of the extant seeds with the SEM was performed in the Key Laboratory for Plant Diversity and Biogeography of East Asia, Kunming Institute of Botany, Chinese Academy of Sciences. Extant seeds of Eurya were provided by the Herbarium of Kunming Institute of Botany, Chinese Academy of Sciences. This study was supported by the National Natural Science Foundation of China (No. U1502231, 31300187). This study is a contribution to NECLIME (Neogene Climate of Eurasia).
| An Z.S., Clemens S.C., Shen J., et al., 2011. Glacial-interglacial Indian summer monsoon dynamics[J]. Science, 333, 719 -723. DOI:10.1126/science.1203752 | ||
| Basilici G., Martinetto E., Pavia G., et al., 1997. Paleoenvironmental evolution in the Pliocene marine-coastal succession of Val Chiusella (Ivrea, NW Italy)[J]. Boll della Soc. Paleontol. Ital, 36, 23 -52. | ||
| Bhandari S., Momohara A., Paudayal K.N., 2009. Late Pleistocene plant macro-fossils from the Gokarna Formation of the Kathmandu Valley, Central Nepal[J]. Bull. Dep. Geol, 12, 75 -88. | ||
| Bhandari S., Paudayal K.N., Momohara A., 2010. Late Pleistocene plant macrofossils from the Thimi Formation (Madhyapur Thimi section) of the Kathmandu Valley, central Nepal[J]. J. Nepal Geol. Soc, 40, 31 -48. | ||
| Boulter M.C., 1971. A survey of the Neogene flora from two derbyshire pocket deposits[J]. Mercian Geol, 4, 45 -62. | ||
| Bozukov V., Palamarev E., 1995. On the Tertiary history of the Theaceae in Bulgaria[J]. Flora Mediterr, 5, 177 -190. | ||
| Bozukov V., Palamarev E., Petkova A., 2008. The fossil macroflora of the Vulche Pole Molasse formation (SE Bulgaria)[J]. Phytol. Balc, 14, 173 -184. | ||
| Chandler, M.E.J., 1963. The Lower Tertiary Floras of Southern England. Ⅱ. Flora of the Bournemouth Beds; the Boscombe, and the Highcliff Sands. British Museum (Natural History), London. | ||
| Dyjor S., Kvacek Z., Łańcucka-.rodoniowa M., et al., 1992. The younger tertiary deposits in the Gozdnica region (SW Poland) in the light of recent palaeobotanical research[J]. Pol. Bot. Stud, 3, 1 -129. | ||
| Ehlers, J., Gibbard, P.L., Hughes, P.D., 2011. Quaternary Glaciations-extent and Chronology. A Closer Look. Elsevier, The Netherlands. | ||
| Friis E.M., 1979. The Damgaard flora: a new Middle Miocene flora from Denmark[J]. Bull. Geol. Soc. Den, 27, 117 -142. | ||
| Friis E.M., 1985. Angiosperm fruits and seeds from the middle Miocene of Jutland (Denmark)[J]. Det. K. Dan. Vidensk. Selsk. Biol. Skr, 24, 1 -165. | ||
| Friis, E.M., Crane, P.R., Pedersen, K.R., 2011. Early Flowers and Angiosperm Evolution. Cambridge University Press, Cambridge. | ||
| Ge, H.R., Li, D.Y., 1999. Cenozoic Coal-bearing Basins and Coal-forming Regularity in West Yunnan. Yunnan Science and Technology Press, Kunming. | ||
| Gregor, H.J., 1982. Die Jungtertiären Floren Süddeutschlands: Paläokarpologie, Phytostratigraphie, Paläoökologie, Paläoklimatologie. Ferdinand Enke Verlag, Stuttgart. | ||
| Knobloch E., 1977. Paläokarpologische Charakteristik der Flyschzone der mährischen Karpaten[J]. Palaeontologie, 19, 79 -135. | ||
| Knobloch, E., Mai, D.H., 1986. Monographie der Früchte und Samen in der Kreide von Mitteleuropa. Rozpravy ústredního ústavu geologickénho, Praha. | ||
| Knobloch E., Mai D.H., 1991. Evolution of Middle and Upper Cretaceous floras in central and western Europe[J]. Geol. Jahrb. Reihe A, 134, 257 -270. | ||
| Li S.H., Deng C.L., Yao H.T., et al., 2013. Magnetostratigraphy of the Dali Basin in Yunnan and implications for late Neogene rotation of the southeast margin of the Tibetan plateau[J]. J. Geophys. Res. Solid Earth, 118, 791 -807. DOI:10.1002/jgrb.50129 | ||
| Łańcucka-Środoniowa M., 1966. Tortonian flora from the "Gdów Bay" in the south of Poland[J]. Acta Palaeobot, 7, 3 -135. | ||
| Łańcucka-Środoniowa M., 1981. Macroscopic plant remains from the Miocene deposits at Stara Wie's near Wilamowice (southern Poland)[J]. Acta Palaeobot, 21, 115 -126. | ||
| Łańcucka-Środoniowa M., 1984. The results obtained hitherto in studies on the Miocene macroflora from the salt-mine at Wieliczka (S. Poland).[J]. Acta Palaeobot, 24, 3 -26. | ||
| Mai D.H., 1960. Über neue Früchte und Samen aus dem deutschen Tertiär[J]. Paläontol. Z, 34, 73 -90. DOI:10.1007/BF02987044 | ||
| Mai D.H., 1971. Über fossile Lauraceae und Theaceae in Mitteleuropa[J]. Feddes Repert, 82, 313 -341. | ||
| Mai D.H., Walther H., 1978. Die Floren der Haselbacher Serie im WeisselsterBecken (Bezirk Leipzig, DDR)[J]. Abh. Staatl. Musems Mineral. Geol. Dresd, 28, 1 -200. | ||
| Martinetto E., 2001. The role of central Italy as a centre of refuge for thermophilous plants in the late Cenozoic[J]. Acta Palaeobot, 41, 299 -319. | ||
| Martinetto, E., Pavia, G., Bertoldi, R., 1997. Fruit and seed floras rich in exotic and subtropical elements from two lower Pliocene successions of Italy. In: Proceedings of the 4th European Palaeobotanical and Palynological Conference, pp. 237-244. Mededelingen Nederlands Instituut voor Toegepaste Geowetenschappen TNO 58. | ||
| Martinetto E., Monegato G., Irace A., et al., 2015. Pliocene and Early Pleistocene carpological records of terrestrial plants from the southern border of the Po Plain (northern Italy)[J]. Rev. Palaeobot. Palynol, 218, 148 -166. DOI:10.1016/j.revpalbo.2014.10.007 | ||
| Meller B., 1998. Diaspore assemblages from the Early Miocene lignite opencast mine Oberdorf (N Voitsberg, Styria, Austria)[J]. Jahrb. Geol. Bundesanst, 140, 453 -460. | ||
| Meller B., Kovar-Eder J., Zetter R., 1999. Lower Miocene leaf, palynomorph, and diaspore assemblages from the base of the lignite-bearing sequence in the opencast mine Oberdorf, N Voitsberg (Styria, Austria) as an indication of “Younger Mastixioid†vegetation[J]. Palaeontogr. Abt. B, 252, 123 -179. | ||
| Min, T.L., Bartholomew, B., 2007. Theaceae, Flora of China. Science Press/Missouri Botanical Garden Press, Beijing/St. Louis. | ||
| Momohara A., 1992. Late Pliocene plant biostratigraphy of the lowermost part of the Osaka group, southwest Japan, with reference to extinction of plants[J]. Quat. Res, 31, 77 -89. DOI:10.4116/jaqua.31.77 | ||
| Momohara A., Saito T., 2001. Change of paleovegetation caused by topographic change in and around a sedimentary basin of the Upper Miocene Tokiguchi Porcelain Clay Formation, central Japan[J]. Geosci. Rept. Shimane Univ. vol, 20, 49 -58. | ||
| Mosbrugger V., Utescher T., Dilcher D.L., 2005. Cenozoic continental climatic evolution of Central Europe[J]. Proc. Natl. Acad. Sci. U.S.A, 102, 14964 -14969. DOI:10.1073/pnas.0505267102 | ||
| Nikitin, V.P., 2006. Palaeocarpology and Stratigraphy of the Paleogene and the Neogene Strata in Asian Russia. Izdatelstvo Akademii "Geo", Novosibirsk. | ||
| Ozaki, K., 1991. Late Miocene and Pliocene Floras in Central Honshu, Japan. Kanagawa Prefectural Museum, Yokohama. | ||
| Shen J., Xiao H.F., Wang S.M., et al., 2007. The orbital scale evolution of regional climate recorded in a long sediment core from Heqing, China[J]. Chin. Sci. Bull, 52, 1813 -1819. DOI:10.1007/s11434-007-0246-5 | ||
| Su T., Jacques F.M.B., Liu Y.S., et al., 2011. A new Drynaria (Polypodiaceae) from the upper Pliocene of southwest China[J]. Rev. Palaeobot. Palynol, 164, 132 -142. DOI:10.1016/j.revpalbo.2010.11.011 | ||
| Tanai T., Uemura K., 1991. The Oligocene Noda flora from the Yuya-wan area of the western end of Honshu, Japan[J]. Part 1. Bull. Natl. Sci. Mus, 17, 57 -80. | ||
| Tao, J.R., 1986. Neogene flora of Lanping and its significance in middle watershed of Selween-Mekong-Yantze Rivers. In: Studies in Qinghai-Xizang (Tibet) Plateau, Special Issue of Hengduan Mountains Scientific Expedition, vol.2. Science and Technology Publishing House, Beijing. http://en.cnki.com.cn/Article_en/CJFDTOTAL-ZWXB198203012.htm | ||
| Tao J.R., Du N.Q., 1982. Neogene flora of Tengchong Basin in western Yunnan, China[J]. Acta Bot. Sin, 24, 273 -281. | ||
| Tao J.R., Kong Z.C., 1973. The fossil florule and sporo-pollen assemblage of the Shang-In coal series of Erhyuan, Yunnan[J]. Acta Bot. Sin, 15, 120 -130. | ||
| Teodoridis V., Kvačcek Z., 2006. Palaeobotanical research of the Early Miocene deposits overlying the main coal seam (Libkovice and Lom members) in the Most Basin (Czech Republic)[J]. Bull. Geosci, 81, 93 -113. | ||
| van der Burgh J., 1978. The Pliocene flora of Fortuna-Garsdorf I[J]. Fruits and seeds of angiosperms. Rev. Palaeobot. Palynol, 26, 173 -211. DOI:10.1016/0034-6667(78)90011-8 | ||
| van der Burgh J., 1983. Allochthonous seed and fruit floras from the Pliocene of the lower Rhine Basin[J]. Rev. Palaeobot. Palynol, 40, 33 -90. DOI:10.1016/0034-6667(83)90004-0 | ||
| van der Burgh J., 1987. Miocene floras in the lower Rhenish basin and their ecological interpretation[J]. Rev. Palaeobot. Palynol, 52, 299 -366. DOI:10.1016/0034-6667(87)90064-9 | ||
| WGVY (Writing Group of Vegetation of Yunnan), 1987. Vegetation of Yunnan. Science and Technology Press, Beijing. | ||
| Wu C.C., Hsu Z.F., Tsou C.H., 2007. Phylogeny and taxonomy of Eurya (Ternstroe-miaceae) from Taiwan, as inferred from ITS sequence data[J]. Bot. Stud, 48, 97 -116. | ||
| Xiao X.Y., Shen J., Wang S.M., et al., 2010. The variation of the southwest monsoon from the high resolution pollen record in Heqing Basin, Yunnan Province, China for the last 2[J]. 78 Ma. Palaeogeogr. Palaeoclimatol. Palaeoecol, 287, 45 -57. DOI:10.1016/j.palaeo.2010.01.013 | ||
| Yamakawa, C., Momohara, A., Saito, T., et al., 2015. Composition and paleoenvironment of wetland forests dominated by Glyptostrobus and Metasequoia in the latest Pliocene (2.6 Ma) in central Japan. Palaeogeogr. Palaeoclimatol. Palae-oecol. (in press). | ||
| Zhu H., Huang Y.J., Ji X.P., et al., 2016. Continuous existence of Zanthoxylum (Rutaceae) in southwest China since the Miocene[J]. Quat. Int, 392, 224 -232. DOI:10.1016/j.quaint.2015.05.020 |