



Coexisting related species are likely to suffer reproductive interference through heterospecific pollen (HP) receipt via pollinator sharing (Morales and Traveset, 2008; Moreira-Hernández and Muchhala, 2019; Zou et al., 2022). Multiple barriers contribute to restricting interspecific gene flow and maintaining species boundaries (Ramsey et al., 2003). According to the order in which they act in the reproductive cycle, reproductive barriers are classified as pre- and post-pollination components (reviewed in Baack et al., 2015). Pre-pollination barriers such as geographic, phenological, and floral barriers play a key role in minimizing interspecific pollen flow (Huang and Shi, 2013; Ma et al., 2016; Paudel et al., 2018; Matsumoto et al., 2021). Post-pollination barriers, which include pollen-pistil incompatibility and hybrid inviability or sterility, can impede the formation and persistence of hybrids after heterospecific pollination (Diaz and Macnair, 1999; Larcombe et al., 2016).
For naturally sympatric species, reproductive isolation (RI) is often positively correlated with genetic distance ('clock-like' accumulation) and pre-pollination barriers are often stronger and contribute more to total RI than post-pollination barriers (Baack et al., 2015). In several cases—Helianthus, Madiinae, Nolana, and Coreopsis—RI increased with increasing genetic distance (Archibald et al., 2005; Jewell et al., 2012; Owens and Rieseberg, 2014). In contrast to long-term sympatric species, geographic isolation plays a key role in allopatric speciation (Tang et al., 2022). On the one hand, the maintenance of species boundaries for historically allopatric taxa often depends on geographic barriers, so other barriers are often weak (Tang et al., 2022). On the other hand, reproductive barriers (especially in the post-pollination stage) between allopatric sister species may be strengthened (Brock, 2009; Knope et al., 2013) because long-term geographical isolation fosters genetic differentiation and species diversification. Until recently, few studies have determined the strength of pre- and post-pollination barriers between historically allopatric sister species (but see Brock, 2009; Tang et al., 2022).
Biotic invasions pose a serious threat to biodiversity (Mack et al., 2000). Recently, anthropogenic introduction has resulted in a loss of geographical barriers between some previously allopatric species and increased their secondary contact (McFarlane and Pemberton, 2019). If multiple reproductive barriers fail to prevent gene flow, the formation of vigorous or maladaptive hybrids between exotic and native congeners may lead to the extinction of native rare species through genetic assimilation or demographic swamping (Levin et al., 1996; Todesco et al., 2016). Therefore, examining the hybridization proneness of exotic species, estimating the synthesis and vigor of experimental hybrids, and evaluating the degree of RI between alien and native species reveals the potential threat from exotic to native species (Knope et al., 2013).
Sagittaria is a genus in Alismataceae that comprises approximately 40 species globally. S. platyphylla and S. montevidensis are native to North and South America, respectively, but invade East, South, and Southeast China through ornamental trade (Wang et al., 2016; 2021). S. montevidensis, introduced to China, is one of the three subspecies, S. montevidensis subsp. montevidensis. S. platyphylla displays powerful clonal propagation and sexual reproduction by the production of stolons, corms, and achenes, leading to its establishment and rapid spread (Adair et al., 2012). Although S. montevidensis has no asexual reproduction, it has a superior reproductive capacity, producing aggregate inflorescence and achenes (Adair et al., 2012; Miguez and Amela García, 2019). The major concern regarding these two invaders originates from Australia and South Africa where they have caused detrimental ecological and economic impacts (Adair et al., 2012; Ndlovu et al., 2020). Hybridization and introgression may represent additional threats to native congeners (Levin et al., 1996; Todesco et al., 2016), because exotic species can accelerate invasion success by hybridization with native congeners (Ainouche et al., 1999; Ellstrand and Schierenbeck, 2006). With the rapid spread of invasive populations and artificial planting, these two invasive and several native Sagittaria species currently have overlapping (sympatric) areas in their species distribution range in East, South, and Southeast China. Particularly, S. platyphylla and S. trifolia (and its subspecies) have secondary contact and share pollinators in microhabitats (Fig. S1). Within the distribution range, most native Sagittaria species are endemic with narrow geographic ranges. S. lichuanensis is listed as endangered on the IUCN Red List of Threatened Species as of 2010. According to the Red List of Biodiversity in China (higher plants volume), S. guayanensis is classified as endangered, while S. lichuanensis and S. potamogetifolia are vulnerable. Considering the vulnerability and rarity of the native species and the highly invasive proneness of the alien species, anthropogenic hybridization experiments between invasive and native Sagittaria species are needed to uncover the potential threat of alien species.
In this study, we examined the strength of four pre-pollination barriers (phenological, floral traits, pollen production, and floral constancy) and four post-pollination barriers (pollen-pistil incompatibility, seed set, seed viability, and seedling survival) between two invasive and five native Sagittaria species. In particular, we aimed to determine if: (1) experimental hybrids could be synthesized, (2) multiple barriers limited gene flow, and (3) RI was correlated with genetic distance. Our objectives were to determine the threat from invasive to native species, shed light on the risk of hybridization between invasive and native congeners, and pose appropriate management suggestions for species conservation.
2. Materials and methods 2.1. Study speciesIn China, Sagittaria comprises seven native species and one subspecies (Wang et al., 2021). In this study, S. guayanensis subsp. lappula (D. Don) Bogin, S. lichuanensis J.K. Chen, X.Z. Sun & H.Q. Wang, S. potamogetifolia Merr., S. pygmaea Miq., S. trifolia L. and S. trifolia subsp. leucopetala (Miq.) Q.F. Wang were selected because they occur in East, South, and Southeast China where exotic S. montevidensis Cham. & Schltdl. and S. platyphylla (Engelm.) J.G. Sm. invade (Wang et al., 2016; 2020). In China, S. trifolia subsp. leucopetala is a cultivated subspecies of S. trifolia with corms for food (Chen, 1989). All experiments were performed with S. trifolia and its subspecies because both were found in the distribution range of two alien species (Fig. S1). Sagittaria species are aquatic perennial or annual herbs, with a flowering period from May to October (Chen, 1989). Most species are monoecious, producing female and male flowers, while S. guayanensis is andromonoecious, producing perfect and male flowers (Chen, 1989; Huang et al., 2000; Huang, 2003; Wang et al., 2021). Sagittaria is protogynous, with female or perfect flowers grown in the proximal whorls of the racemose or paniculate inflorescence and bloom first, while male flowers in the distal whorls of the inflorescence bloom later (Chen, 1989; Adair et al., 2012). The bowl-shaped flower comprises three green sepals and three white petals (Chen, 1989). For S. guayanensis and S. montevidensis subsp. montevidensis, the base of the petal often has a purple or dark reddish spot (Fig. S2; Huang, 2003; Adair et al., 2012; Miguez and Amela García, 2019). The female and perfect flower bear numerous apocarpous carpels which are dense on the globose receptacle and have a single ovule per carpel (Chen, 1989; Adair et al., 2012; Wang et al., 2021). The achene has a beak formed by the style and its seeds appear horseshoe-shaped with brown testa (Chen, 1989; Adair et al., 2012). Sagittaria is mainly pollinated by generalized pollinators like flies and bees (Huang et al., 2006; Adair et al., 2012; Miguez and Amela García, 2019). Some species (except S. guayanensis and S. montevidensis) can expand their populations by producing abundant ramets from corms, bulbils, and stolons (Chen, 1989; Adair et al., 2012). Species photos are provided in Fig. S2.
2.2. Study siteHundreds of wild individuals for each Sagittaria species (except S. trifolia subsp. leucopetala and S. montevidensis) were naturally flowering in the field population (Table S1). Thirty individuals of each species were randomly collected and transplanted to the Garden of Greenhouse within Wuhan University (30°32′N, 114°21′E), Wuhan, Hubei Province, China at the end of May 2021 and 2022. When collecting species with clonal ability, the distance between focal individuals was more than 2 m to avoid sampling ramets. At the same time, 30 individuals of cultivated S. trifolia subsp. leucopetala and S. montevidensis were sampled from Tianjing Aquatic Nursery, Hangzhou, China (Table S1). A single individual was placed in a 9.7 L or 50 L plastic pot filled with loam soil according to the size of the plant. Normal water and fertilizer management was carried out regularly, and light and temperature were provided naturally. To avoid potential hybrid genetic pollution to parental plants, all fruits produced by open pollination were removed before maturation. Two pre-pollination barriers (phenological isolation and floral traits) were estimated in field populations in July 2022, whereas all other barriers were evaluated in the Garden of Greenhouse within Wuhan University from June to September in 2021 and 2022. For S. trifolia subsp. leucopetala and S. montevidensis, all experiments were conducted in the greenhouse within Wuhan University in 2021 and 2022.
2.3. Pre-pollination barriers 2.3.1. Flowering phenology and pollen viabilityThe seasonal flowering phenology of all species completely overlapped from the end of May to the beginning of October (Zou Ting–Ting, personal observation). Therefore, the strength of the phenological barrier was assessed based on the functional daily flowering time associated with pollinator activity and pollen viability in July 2022. A total of 50 male flowers and 50 female flowers (perfect flowers of Sagittaria guayanensis) were randomly selected from 25 individuals of each species. Open flowers were counted hourly from 07:00 to 17:00. At the same time, the viability of the pollen mixture of another 20 male flowers or perfect flowers from these individuals was detected with 1% (w/v) 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) under a light microscope (Nikon E100; Nikon Corp., Chiyoda, Japan) every 1 h or 2 h (Qiu et al., 2021). According to the degree of dyeing, pollen viability was divided into three groups: 100% = dark purple; 50% = light purple; and 0% = colorless. Phenological observations were conducted on three sunny days for each species.
2.3.2. Floral traitsTwenty-five individuals from each species were randomly selected and the differences in floral morphology among species were evaluated. Traits of two male and two female flowers (perfect flowers of Sagittaria guayanensis) per individual were measured with a digital caliper: (1) pedicel length, (2) corolla diameter, (3) petal length, (4) petal width, (5) gynoecium/androecium diameter, (6) stamen number, and (7) number of spots on petals per flower.
2.3.3. Pollen productionPollen production is closely correlated with pollinator preference and constancy, reflecting the proportion of HP and conspecific pollen (CP) within the pollen pool transported to the ovule (Brys et al., 2016). In 2021, the total number of inflorescences of 9–12 individuals was counted for each species. From May to July 2022, 50 inflorescences from 30 individuals were randomly selected for each species, and the total number of male flowers (and perfect flowers of S. guayanensis) per inflorescence was counted. Owing to the limited number of inflorescences, 21 inflorescences from 30 individuals were randomly labeled, and the total number of male flowers was counted for S. trifolia subsp. leucopetala. For these 30 individuals, 30 male flowers (and perfect flowers of S. guayanensis) were randomly collected, and the pollen production per flower was determined. The flower buds were collected the day before anthesis and fixed in FAA solution (70% (v/v) alcohol: acetic acid: formalin, 90:5:5). The mature undehisced anthers were removed with tweezers, crushed by a grinding rod to release pollen grains, and transferred into slightly less than 1000 μl of water. The accurate volume of the pollen stoste was estimated using the 1000-μl pipette (Eppendorf GmbH, Hamburg, Germany) by rotating the knob to exhaust air. The pollen stoste was then diluted to the appropriate concentration and the pollen grains contained in a 5-μl diluted drop were counted under a light microscope (Nikon E100; Nikon Corp., Chiyoda, Japan). Counts were averaged for three replicates and recalculated as the pollen production per male/perfect flower based on the dilutability and stoste volume. Mean pollen production per individual was calculated according to mean pollen production of a single flower, mean flower number per inflorescence, and mean inflorescence number per individual.
2.3.4. Pollinator foraging behaviorPreliminary work showed that Sagittaria flowers were predominantly visited by Apis cerana, Halictus pulvereus, Andrena camellia, some flies, and some butterflies both in the field and greenhouse (Zou Ting–Ting, unpublished data). Artificial arrays comprising exotic and native species pairs were established to estimate floral constancy in the Garden of Greenhouse within Wuhan University. To eliminate the impact of floral display on pollinator attraction to Sagittaria (Huang et al., 2006), each array comprised two rows of pots, one with five pots of exotic species and the other with five pots of native species. For each pot, one female flower (perfect flowers of S. guayanensis) and one male flower from different inflorescences were retained. Thus, each array consisted of five male and five female/perfect flowers from exotic and native species. Foam boxes were used to raise the height of the shorter species so that the soil level height of the two species was consistent. On the same day, at most two arrays 30 m apart from the same species pair were established. For each species pair, five or six arrays were established during overlapping anthesis. Before observation, the buds of other Sagittaria plants in the garden were removed to avoid irrelevant pollen and floral scents interfering with the interaction between formulated pairs. Visitors that made contact with gynoecium or androecium and carried pollen were identified as legitimate pollinators. For each foraging bout in the array, the pollinated species, visitation sequence to the flowers, and flower sex were recorded from 7:00 to 14:00 on sunny days. Each observation period of an experimental array lasted for 15 min. If only one flower was visited by pollinators in a foraging bout, then that bout was regarded as an occasional visit and deleted from subsequent analyses (Tong and Huang, 2016). The floral constancy index and pollinator preference index were calculated using the equation from Castro et al. (2020, Table S2). These indices ranged from 0 to 1, where a value of 1 indicated complete constancy with a species and preference for one species, while a value of 0.5 indicated no constancy or preference.
2.4. Post-pollination barriers 2.4.1. Pollen-pistil barriersArtificial reciprocal pollination was performed to evaluate the strength of the pollen-pistil barrier. Eight intraspecific crosses and 24 interspecific crosses between exotic and native species were performed. Eight to fourteen maternal individuals were randomly selected, and one female flower (perfect flower of Sagittaria guayanensis) per individual was randomly labeled in each cross. These female/perfect and male flower buds of both exotic and native species were bagged with mesh netting before flowering. In particular, the perfect flowers of S. guayanensis were previously emasculated to prevent the interference of pre-anthesis cleistogamy (Wang et al., 2006). For each exotic-native species pair, either CP or HP was deposited directly on the stigma to saturation at the beginning of overlapping flowering to ensure maximum stigma viability and pollen receptivity. The pollinated flowers were re-bagged to exclude pollinators, harvested at 5 h after pollination, fixed with Carnoy's fluid (100% alcohol: acetic acid, 3:1) for 1 h, and then stored in FAA solution. Pollen germination, pollen tube growth, and ovule penetration were observed under a fluorescence microscope (Olympus BX43; Olympus Co., Tokyo, Japan) after treatment with 20% NaOH for 5 h and 0.1% aniline blue for 1 h. Approximately 50 pistils per flower were randomly observed, and 17, 456 pistils were examined for all crosses. Given that Sagittaria has only a single ovule per pistil (Chen, 1989; Adair et al., 2012), the percentage of ovule penetration was calculated as the ratio between the number of pistils with penetrated ovules and the total number of pistils recorded for each flower.
2.4.2. Seed setUsing the same pollination methods as in Section 2.4.1, reciprocal pollination was conducted between exotic and native species to detect hybrid seed formation. Thirty female or perfect flowers from 30 individuals were processed in each cross. The mature aggregate fruits were harvested about two weeks later and dried for 5 h in an electric heating constant temperature drying box (GZX-9030 MBE, Boxun Co., Shanghai, China) at 50 ℃. The achenes of each fruit were dissected and divided into three types (undeveloped, partly developed, and fully developed) according to achene size and the absence or presence of an embryo (Huang et al., 2000; Huang, 2003) under a stereoscope (Olympus SZX16, Olympus Co., Tokyo, Japan). Approximately 100 achenes per aggregate fruit were randomly observed, and 115, 362 achenes from 960 aggregate fruits were examined for all crosses. Since one pistil produced only one ovule (Chen, 1989; Adair et al., 2012), seed formation was equal to achene formation. The seed set was calculated as the ratio between the number of fully developed achenes and the total number of achenes.
Molecular validation of the putative hybrid seeds was conducted using a nuclear internal transcribed spacer (ITS). For each interspecific cross that produced fully developed seeds, 5–8 seeds from three different parental combinations were randomly selected. Total genomic DNA for parent species was extracted from leaf tissue using Hi-DNAsecure Plant Kit (DP350, Tiangen Biotech Co., Ltd., Beijing, China), while genomic DNA from seeds was extracted from the micro embryo using Chelex100 chelating resin according to the method provided in Walsh et al. (1991). PCR was conducted to amplify the ITS sequence using primers ITS-4 and ITS-5 (Baldwin, 1992) on a PCR Amplifier (Eppendorf GmbH, Hamburg, Germany). The PCR program was as follows: 94 ℃ (4 min), 30 cycles of 94 ℃ (30 s), 53 ℃ (40 s), 72 ℃ (30 s), and a final extension of 72 ℃ (10 min). PCR products were purified using DNA Gel Extraction Kit (TSP 601, Tsingke Biotechnology Co., Ltd., Beijing, China), attached to plasmids using pClone007 Versatile Simple Vector Kit (TVS-007VS, Tsingke Biotechnology Co., Ltd., Beijing, China), and then transformed into Trelief™ 5ɑ Chemically Competent Cell (TSC-C01, Tsingke Biotechnology Co., Ltd., Beijing, China). About 10 positive clones for parent species and about five clones for seeds were randomly selected and sequenced by Huayu Gene Biotechnology Co., Ltd. (Wuhan, China) using M13F and M13R primers (Table S3). All newly generated sequences were submitted to figshare database (Zou, 2022). These sequences were aligned using Geneious Prime 2021 (Biomatters, Auckland, New Zealand), and polymorphic sites and haplotypes were analyzed using DNASP 6 (Librado and Rozas, 2009; Rozas et al., 2017). Finally, haplotype networks were formed using median-joining methods by PopART 1.7 (Bandelt et al., 1999).
2.4.3. Seed viability and seedling survivalGiven that preliminary work showed that partly developed achenes (even comprising half the embryo) are inviable and fail to germinate (Zou Ting–Ting, unpublished data), the germination experiment was conducted in fully developed achenes to estimate the strength of seed viability and the seedling survival barrier. Reciprocal pollination was performed between exotic and native species using the same pollination method as in Sections 2.4.1 and 2.4.2. For each cross, 20 mature aggregate fruits were collected from 20 maternal individuals and stored dry at 4 ℃ for four months. One hundred fully developed achenes were selected under a stereoscope (Olympus SZX16, Olympus Co., Tokyo, Japan) and placed on a 5.5 cm plastic petri dish with two filter papers moistened with distilled water. Petri dishes were sealed with parafilm to minimize water loss and then placed in growth chambers (GZC-300C; Youke Instrument Equipment Co., Ltd., Hefei, China; day/night, 14/10 h, 33/20 ℃, 16660/0 lux). The number of germinated seeds was counted using visible radicle protrusions as a standard to determine germination success every week. At the same time, those seedlings with normal morphology were placed in a new Petri dish and the survival number was scored using green leaves as a standard the next week. Seedling survival was estimated as a proxy for early age barriers to F1 hybrid success. Seed germination ceased eight months later. The seed germination rate and seedling survival rate of each culture dish were calculated according to the total number of germinated seeds and surviving seedlings.
2.5. Genetic distanceThe pairwise genetic distance between exotic and native species was calculated for each exotic-native species pair using four DNA regions: ITS, psbA-trnH, matK, and rbcL. All sequences were obtained from GenBank (Table S4). A representative from Sagittaria trifolia was substituted for S. trifolia subsp. leucopetala because the exact sequences could not be found. Given that S. trifolia subsp. leucopetala is a subspecies, it was unlikely to influence the results. Alignments were performed on each separate DNA region, and then those DNA regions were concatenated into a super matrix in Geneious Prime 8.0.4 (Biomatters, Auckland, New Zealand). Finally, the pairwise genetic distances were calculated using MEGA 11.0 (Tamura et al., 2021).
2.6. Statistical analysisTemporal isolation (flowering phenology) was calculated using the equation RI4C by Sobel and Chen (2014) (Table S5). The strength of the pollen production barrier, pollinator-mediated isolation, pollen-pistil barrier, seed set, seed viability, and seedling survival was calculated following the RI4A equation by Sobel and Chen (2014) (Table S5). The total RI was calculated based on equation RI4E by Sobel and Chen (2014) (Table S5). The relative and absolute contributions of individual barriers to total isolation were also calculated following the equations by Sobel and Chen (2014) (Table S5).
A principal component analysis (PCA) was conducted to detect the degree of morphological overlap between alien and native species. After normality testing, the differences in pollen production between native and alien species were analyzed using one-way ANOVA, and significant differences in the post-hoc multiple comparison were identified using the Games-Howell method. A Pearson Chi-squared test was used to examine the deviations from 0.5 (no floral constancy or pollinator preference) for these two indices for each species pair. One-way ANOVA adapted with the Games-Howell post hoc test or Kruskal–Wallis test adapted with the Dunn post hoc test was used to detect the percentage of ovule penetration difference between different crosses. The differences in seed set and seed germination between different species combinations were identified using one-way ANOVA with the Games-Howell method in the post-hoc multiple comparison. A Kruskal–Wallis test adapted with Dunn's post hoc tests was used to determine seedling survival differences between different species pairs. Spearman rank correlation analysis was conducted to examine the effects of genetic distance on the individual barrier (except for seed viability and seedling viability). All data analyses were performed using SPSS 28.0 (IBM Co., Armonk, NY, USA). A P-value of ≤0.05 was considered statistically significant.
3. Results 3.1. Pre-pollination barriers 3.1.1. Flowering phenologyDaily flowering phenology of exotic and native species highly overlapped (Fig. 1). RI was therefore low and varied among species pairs (RI range: 0–0.5000, Table 1).

|
| Fig. 1 Daily flowering phenology of two exotic and five native (including one subspecies) Sagittaria species. Shaded bars show the mean number of female (perfect flowers for S. guayanensis) and male flowers at anthesis. Circles and error bars represent the mean percentage of viable pollen and standard errors. |
| Maternal × Paternal | Strength of RI | Total RI | ||||||
| Phenology | Pollen production | Floral constancy | Pollen-pistil incompatibility | Seed set | Seed viability | Seedling viability | ||
| S. pygmaea × S. platyphylla | 0.0000 | −0.9220 | 0.2113 | 0.0326 | 1.0000 | – | – | 1.0000 |
| S. platyphylla × S. pygmaea | 0.2500 | 0.9220 | 0.8210 | 0.0740 | 0.1914 | 0.5761 | 0.8815 | 0.9999 |
| S. trifolia × S. platyphylla | 0.1000 | −0.0720 | 0.6332 | 0.0495 | 0.0590 | 0.1669 | 0.0033 | 0.7697 |
| S. platyphylla × S. trifolia | 0.0000 | 0.0720 | 0.3600 | 0.0695 | 0.0504 | 0.2742 | 0.0778 | 0.7299 |
| S. trifolia subsp. leucopetala × S. platyphylla | 0.1000 | −0.1419 | 0.8323 | 0.1678 | 0.0865 | 0.0375 | 0.0093 | 0.8880 |
| S. platyphylla × S. trifolia subsp. leucopetala | 0.0000 | 0.1419 | 0.5636 | −0.0117 | 0.0413 | 0.2579 | 0.1811 | 0.8504 |
| S. potamogetifolia × S. platyphylla | 0.0000 | −0.5745 | 0.4930 | 0.0769 | 1.0000 | – | – | 1.0000 |
| S. platyphylla × S. potamogetifolia | 0.3750 | 0.5745 | 0.5823 | 0.0835 | 1.0000 | – | – | 1.0000 |
| S. lichuanensis × S. platyphylla | 0.0000 | −0.4533 | 0.5072 | −0.4609 | 1.0000 | – | – | 1.0000 |
| S. platyphylla × S. lichuanensis | 0.3750 | 0.4533 | 0.6901 | 0.7044 | 1.0000 | – | – | 1.0000 |
| S. guayanensis × S. platyphylla | 0.0000 | −0.8925 | 0.3333 | 0.0665 | 1.0000 | – | – | 1.0000 |
| S. platyphylla × S. guayanensis | 0.5000 | 0.8925 | 0.7725 | 0.2325 | 1.0000 | – | – | 1.0000 |
| S. pygmaea × S. montevidensis | 0.3333 | −0.9843 | 0.4857 | 0.0160 | 1.0000 | – | – | 1.0000 |
| S. montevidensis × S. pygmaea | 0.2857 | 0.9843 | 0.9262 | 0.8659 | 1.0000 | – | – | 1.0000 |
| S. trifolia × S. montevidensis | 0.2000 | −0.7110 | 0.6822 | 0.0671 | 1.0000 | – | – | 1.0000 |
| S. montevidensis × S. trifolia | 0.0000 | 0.7110 | 0.6696 | 0.2322 | 1.0000 | – | – | 1.0000 |
| S. trifolia subsp. leucopetala × S. montevidensis | 0.2000 | −0.7443 | 0.3445 | 0.0344 | 1.0000 | – | – | 1.0000 |
| S. montevidensis × S. trifolia subsp. leucopetala | 0.0000 | 0.7443 | 0.6530 | 0.1996 | 1.0000 | – | – | 1.0000 |
| S. potamogetifolia × S. montevidensis | 0.3333 | −0.8998 | 0.3973 | 0.7162 | 1.0000 | – | – | 1.0000 |
| S. montevidensis × S. potamogetifolia | 0.4286 | 0.8998 | 0.8732 | 0.9886 | 1.0000 | – | – | 1.0000 |
| S. lichuanensis × S. montevidensis | 0.3333 | −0.8633 | 0.3704 | −0.5987 | 1.0000 | – | – | 1.0000 |
| S. montevidensis × S. lichuanensis | 0.4286 | 0.8633 | 0.7455 | 0.9551 | 1.0000 | – | – | 1.0000 |
| S. guayanensis × S. montevidensis | 0.0000 | −0.9781 | 0.6327 | −0.0487 | 1.0000 | – | – | 1.0000 |
| S. montevidensis × S. guayanensis | 0.4286 | 0.9781 | 0.9361 | 0.7785 | 1.0000 | – | – | 1.0000 |
For the PCA, principal component axes 1 and 2 explained 60.53% and 16.70% of the total variation of female floral traits and 39.08% and 16.78% for male floral traits, respectively. For both female and male flowers, all morphological measures contributed to the variation of principal component axis 1. Sagittaria montevidensis had the largest floral display and showed no overlap in floral traits with any native species (Fig. 2). Floral traits differed considerably between S. guayanensis and the two alien species (Fig. 2). However, S. platyphylla floral phenotypes overlapped with all native species (except for S. guayanensis, Fig. 2).

|
| Fig. 2 Principal component analysis of floral morphology for two exotic and five native (including one subspecies) Sagittaria species. (A) Female flowers (perfect flowers for S. guayanensis). (B) Male flowers. |
The four components of pollen production differed significantly between species (Table 2). Although the pollen production per flower and the male flower number per inflorescence of the two alien species was not significantly higher than that of all native species, their inflorescence number per individual was significantly higher than that of all native species (F7,81 = 118.023, P < 0.001, Table 2). As a result, alien species produced significantly more pollen grains per individual than did native species (Table 2), resulting in asymmetric RI within species pairs and stronger RI in alien species (RI range: −0.9843 to 0.9843, Table 1).
| Species | Number of pollen grains per flower | Number of male flowers per inflorescence | Number of inflorescences per individual | Total number of pollen grains per individual |
| S. pygmaea | 2.5 × 104 ± 6.0 × 103a (30) | 3.9 ± 0.6a (50) | 14.8 ± 1.9a (12) | 1.4 × 106 |
| S. trifolia | 9.9 × 104 ± 2.6 × 104b (30) | 36.3 ± 12.9b (50) | 8.6 ± 1.4b (10) | 3.1 × 107 |
| S. trifolia subsp. leucopetala | 1.8 × 105 ± 4.1 × 104c (30) | 82.2 ± 22.0c (21) | 1.8 ± 1.0c (10) | 2.7 × 107 |
| S. potamogetifolia | 5.4 × 104 ± 1.4 × 104d (30) | 9.0 ± 2.0d (50) | 19.8 ± 3.1d (12) | 9.6 × 106 |
| S. lichuanensis | 9.7 × 104 ± 3.1 × 104b (30) | 26.7 ± 8.4e (50) | 5.2 ± 1.5e (12) | 1.3 × 107 |
| S. guayanensis (male) | 2.9 × 104 ± 6.8 × 103a (30) | 3.0 ± 1.0f (50) | 18.3 ± 2.7 ad (9) | 2.0 × 106 |
| S. guayanensis (perfect) | 8.6 × 103 ± 2.5 × 103e (30) | 2.8 ± 0.7f (50) | ||
| S. platyphylla | 5.8 × 104 ± 1.8 × 104d (30) | 8.5 ± 2.0d (50) | 72.2 ± 16.6f (12) | 3.6 × 107 |
| S. montevidensis | 3.4 × 105 ± 7.6 × 104f (30) | 18.6 ± 4g (50) | 28.8 ± 5.6g (12) | 1.8 × 108 |
For each species pair, pollinator visitation was observed for 225–765 min (Table 3). Pollinator visit rates were significantly higher for alien species than for native species for most species pairs, indicating a significant pollinator preference for exotic species (except for Sagittaria platyphylla-S. trifolia and S. platyphylla-S. trifolia subsp. leucopetala; Table 3, Fig. S3). Although most pollinators visited both alien and native species when foraging in the quadrats, interspecific visits occurred infrequently, leading to a significantly high floral constancy index (Table 3) and low interspecific pollen transfer (Fig. S4). Different generalized pollinators showed different floral constancy index values (Fig. S5). High pollinator preference and floral constancy resulted in asymmetric RI within species pairs and stronger RI in alien species (Table 1).
| Species pair (a vs. b) | Time (min) | No. of floral visits | CI | PI | |||||||
| a → a | b → b | a → b | b → a | mean | P | mean | P | ||||
| S. platyphylla vs S. pygmaea | 765 | 234 | 43 | 28 | 23 | 0.845 | < 0.001 | 0.770 | < 0.001 | ||
| S. platyphylla vs S. trifolia | 465 | 153 | 285 | 64 | 72 | 0.763 | < 0.001 | 0.313 | < 0.001 | ||
| S. platyphylla vs S. trifolia subsp. leucopetala | 300 | 43 | 142 | 13 | 12 | 0.881 | < 0.001 | 0.295 | < 0.001 | ||
| S. platyphylla vs S. potamogetifolia | 375 | 125 | 106 | 36 | 33 | 0.770 | < 0.001 | 0.539 | 0.263 | ||
| S. platyphylla vs S. lichuanensis | 360 | 120 | 104 | 34 | 22 | 0.800 | < 0.001 | 0.537 | 0.294 | ||
| S. platyphylla vs S. guayanensis | 510 | 187 | 34 | 17 | 24 | 0.844 | < 0.001 | 0.788 | < 0.001 | ||
| S. montevidensis vs S. pygmaea | 360 | 261 | 26 | 9 | 10 | 0.938 | < 0.001 | 0.869 | < 0.001 | ||
| S. montevidensis vs S. trifolia | 390 | 192 | 217 | 41 | 38 | 0.838 | < 0.001 | 0.525 | 0.360 | ||
| S. montevidensis vs S. trifolia subsp. leucopetala | 225 | 181 | 80 | 39 | 38 | 0.772 | < 0.001 | 0.663 | < 0.001 | ||
| S. montevidensis vs S. potamogetifolia | 315 | 325 | 51 | 22 | 22 | 0.895 | < 0.001 | 0.822 | < 0.001 | ||
| S. montevidensis vs S. lichuanensis | 225 | 96 | 37 | 17 | 14 | 0.811 | < 0.001 | 0.681 | < 0.001 | ||
| S. montevidensis vs S. guayanensis | 480 | 303 | 40 | 9 | 10 | 0.948 | < 0.001 | 0.868 | < 0.001 | ||
Only S. montevidensis showed incongruous pollen tube growth of HP at 5 h post pollination, whereas other species showed congruous ovule penetration for HP (Table 4; Fig. S6). Consequently, barrier strengths were highly variable and asymmetric for each species combination, ranging from −0.5987 to 0.9886.
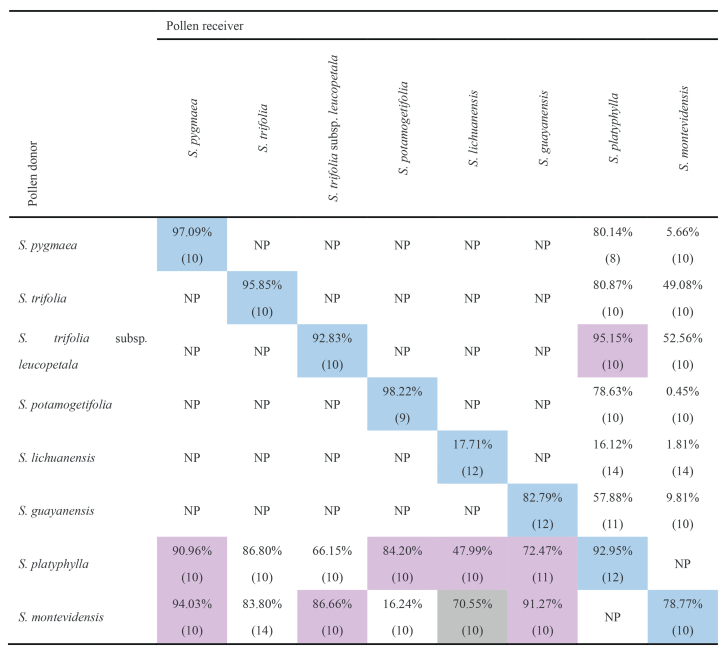
|
Among all the interspecific crosses, only Sagittaria platyphylla successfully produced numerous well-developed seeds with native species (S. pygmaea, S. trifolia, and S. trifolia subsp. leucopetala; Fig. 3 and Table S6). Haplotype networks showed that S. platyphylla and native species (S. pygmaea and S. trifolia) did not share haplotypes with each other. All putative hybrid seeds possessed both maternal and paternal haplotypes (Fig. 4). This molecular evidence indicated that anthropogenic hybridization occurred. Although the seed set was significantly higher from the intraspecific cross than interspecific hybridization for S. trifolia, S. trifolia subsp. leucopetala, and S. platyphylla (F7,232 = 49.913, P < 0.001, Fig. 3A), the barrier in these species combinations was low (RI range: 0.0413–0.1914, Table 1). In contrast, S. montevidensis was incapable of hybridizing with any native species, resulting in RI = 1 (Tables 1 and S6).

|
| Fig. 3 Percentage of seed set, seed germination, and seedling survival. (A) Seed set. (B) Seed germination. (C) Seedling survival. Data of seed set, and seed germination are represented as mean ± SE, while data of seedling survival is showed as median. Broken lines separate crosses with different maternal species. Values with the same lowercase letters are not significantly different (P > 0.05). S. tri, Sagittaria trifolia; S. leu, S. trifolia subsp. leucopetala; S. pla, S. platyphylla; S. pyg, S. pygmaea. |

|
| Fig. 4 Haplotype networks inferred from nuclear internal transcribed spacers for exotic Sagittaria platyphylla, two native congeners, and artificial hybrids. (A) S. trifolia × S. platyphylla. (B) S. platyphylla × S. pygmaea. (C) S. platyphylla × S. trifolia. Mutation steps are indicated, while single mutations are not shown. |
Seed germination success and seedling survival success were significantly different among different crosses (F7,152 = 62.254, P < 0.001, Fig. 3B; H = 125.467, P < 0.001, Fig. 3C). For S. trifolia, conspecific seeds exhibited significantly higher germination success than hybrid seeds, while survival success was not significantly different between hybrid and conspecific seeds (Fig. 3B and C). For S. platyphylla, hybrid seeds had significantly lower germination success and survival success than conspecific seeds (Fig. 3B and C). In contrast, seed germination success and seedling survival success were not significantly different between interspecific hybridization and intraspecific crossing for S. trifolia subsp. leucopetala (Fig. 3B and C). Therefore, the RI was low and varied among species pairs (seed viability, RI range: 0.0375–0.5761, seedling survival, RI range: 0.0093–0.8815, Table 1).
3.3. Total RI and relative contribution of individual barriersRegarding pre-pollination barriers, pollen production and floral constancy had higher RI than phenology in exotic species, whereas floral constancy had the highest RI in native species (Table 1). For post-pollination barriers, the highest RI was found for seed sets in both alien and native species, and the pollen-pistil barrier had a high RI in Sagittaria montevidensis (Table 1). The total RI of most species pairs reached its highest value (total RI = 1). However, for two alien-native species pairs (S. platyphylla-S. trifolia and S. platyphylla-S. trifolia subsp. leucopetala), combining the effects of all barriers resulted in low total RI values in reciprocal crosses (total RI range: 0.7299–0.8880, Table 1). Although S. platyphylla produced hybrid seeds following pollination by S. pygmaea, total RI still had a high value of RI = 0.9999. Despite the high RI value of post-pollination barriers, the relative contributions of pre-pollination barriers particularly that of pollen production, were stronger in two exotic plants (Table S7). In native plants, the only post-pollination barrier to make a relatively large contribution to total RI was failure to seed set. However, when S. trifolia and S. trifolia subsp. leucopetala were paired with S. platyphylla, pre-pollination barriers (especially pollinator constancy) contributed considerably to total RI (Table S7).
3.4. Relationships between genetic distance and RIFor exotic and native Sagittaria species pairs, pairwise genetic distance was not significantly associated with phenological isolation (r = 0.387, P = 0.062), pollen production isolation (r = 0.000, P = 1.000), floral constancy isolation (r = 0.103, P = 0.631), or pollen-pistil incompatibility isolation (r = 0.282, P = 0.182), but was positively correlated with seed set isolation (r = 0.702, P < 0.001, Fig. 5).

|
| Fig. 5 Relationship between genetic distance and different reproductive isolation barriers. Isolation barriers include (A) flowering phenology, (B) pollen production, (C) floral constancy, (D) pollen-pistil incompatibility, and (E) seed set. P-value < 0.05 is in bold. |
Recently, there have been reports of hybridization between invasive plants and native congeners, which pose a threat to native genetic integrity and species boundaries (Ayres et al., 1999; Burgess et al., 2008; Kellner et al., 2012; Zaya et al., 2015). Thus, estimating the relative importance of different pre- and post-pollination barriers to gene flow between related species is important to reveal the potential threat from alien species. In this study, we assessed four pre- and post-pollination barriers and their relative contribution to total RI in several invasive and native Sagittaria species. Results showed a combination of pre- and post-pollination barriers that contributed to prevent interspecific gene flow between alien and native species (except for S. platyphylla-S. trifolia and S. platyphylla-S. trifolia subsp. leucopetala pair). We also demonstrated that the two introduced species had significantly different pre- and post-pollination interactions with native species. However, it is important to acknowledge that our ability to detect real RI may be limited even though we conducted 12 alien–native cross combinations in a greenhouse. To our knowledge, this is the first study to assess the role of both pre- and post-pollination barriers in alien and native sister species. It is possible that RI may be significantly different at a zone of secondary contact, and future studies should evaluate the role of different barriers in maintaining species boundaries using a larger set of exotic-native species in a sympatric distribution area.
Reproductive barriers between alien and native Sagittaria species were highly asymmetric, which is consistent with the results of previous studies on long-term sympatric species (Lowry et al., 2008; Brys et al., 2014; Larcombe et al., 2016). Such asymmetry tends to originate from interspecific divergence in floral morphology, floral display, pollen production, and mating system (Brys et al., 2014; Minnaar et al., 2019; Munguía-Rosas and Jácome-Flores, 2020). Asymmetry also existed in the seed set between S. platyphylla and S. pygmaea. When S. pygmaea served as the pollen parent and S. platyphylla as the seed parent, the cross was highly successful, but the reciprocal cross failed. In contrast, the total RI was symmetric and incomplete between S. platyphylla and S. trifolia (and its subspecies). Hybridization may lead to introgression because F1 hybrids may act as a bridge for bi-directional gene flow between parental species (Larcombe et al., 2016). To our knowledge, this is the first experiment to show that one species and its subspecies have similar interaction patterns as exotic species, although they have different traits.
In all reproductive barriers, the role of floral isolation mediated by generalist pollinators remains controversial, because generalist pollinators often lead to HP deposition (Morales and Traveset, 2008; Moreira-Hernández and Muchhala, 2019). However, pollinator constancy increased total RI between alien and native Sagittaria species, which corroborates previous studies that even generalist pollinators contribute to maintaining species integrity (Keller et al., 2021). The pollinator preference—the result of larger floral display and higher pollen production—may directly lead to strong pollinator constancy in two alien species. Future studies should focus on the contribution of such generalized pollinators to species integrity and pollinator discrimination between sister lineages.
Sympatric taxa tend to evolve stronger pre-pollination barriers to maintain species stability because the cost at the post-pollination stage may be higher (Lowry et al., 2008; Brys et al., 2014; 2016). By contrast, previous research has indicated that post-pollination barriers between newly sympatric and historical allopatric congeners contribute more to total RI (Brock, 2009; Knope et al., 2013). Our results also showed that compared to pre-pollination barriers, post-pollination barriers (e.g., hybrid seed set failure) contributed more to limiting gene flow from exotic congeners to native Sagittaria species. However, we cannot ignore the fact that pre-pollination barriers played a leading role in maintaining species boundaries for two exotic species despite a high value for a post-pollination barrier (reduced seed set). Multiple barriers act in hierarchical order, and thus early pre-pollination barriers can contribute more to total RI than late strong post-pollination barriers (Dell'olivo et al., 2011; Ma et al., 2016). For two introduced Sagittaria species, the strong pre-pollination barriers may be attributed to the significantly larger floral display, higher pollen production, and thus higher pollinator constancy. Our results were consistent with Brock (2009), which showed that pre-pollination barriers are not always redundant upon first contact of alien and native species.
In this study, the pollen-pistil barrier was asymmetric in alien-native species pairs and had a higher RI value in alien species. Such pollen tube incompatibility is an important and common mechanism for avoiding hybrid formation and impeding interspecific gene flow in angiosperms (Matallana et al., 2016; Tong and Huang, 2016). For most flowering plants, inharmonious pollen-tube growth is reflected in the inhibition of HP germination on the stigma and the suppression of pollen tube growth along the style (Martin, 1970; Diaz and Macnair, 1999; Brock, 2009). In addition to these, Sagittaria with extra-gynoecial compitum exhibited another special incongruity phenomenon: heterospecific pollen tubes in some species combinations first passed through the carpel base and turned to the receptacle rather than directly entering the ovules (Zou Ting–Ting, unpublished data), as reported between sympatric S. pygmaea and S. trifolia (Lyu et al., 2016). Future studies should focus on the potential molecular and cellular processes driving this unique pollen-pistil interaction. Furthermore, both native and exotic Sagittaria species displayed CP-tube growth advantage in which pollen tubes of CP entered the ovary earlier than those of HP (Fig. S6). CP-tube advantage is recognized as a mechanism to preserve the ovules for conspecific fertilization (Diaz and Macnair, 1999; Zhong et al., 2019), but CP-tube advantage may not always result in conspecific seed-siring advantage due to pollen attrition, pollen-pistil interactions, and male-male interactions (Bertin, 1990; Carney et al., 1996; Montgomery et al., 2010). Thus, we did not consider the CP-tube advantage as a barrier in this study. Nevertheless, we still highlight the importance of assessing the role of CP-tube advantage in maintaining species integrity for alien and native Sagittaria species following mixed pollination treatments.
Reproductive barriers are often positively correlated with genetic distance in sympatric species (Archibald et al., 2005; Jewell et al., 2012; Owens and Rieseberg, 2014), which is called "clock-like" accumulation of RI (Baack et al., 2015). In this study, we found clear evidence that seed set barrier was the only stage that had a positive relationship with genetic distance between exotic and native species. Only Sagittaria platyphylla-S. trifolia and S. platyphylla-S. pygmaea produced hybrid seeds, which had the shortest pairwise genetic distance among all species combinations. This is potentially the first direct experimental evidence that at least one barrier evolved with divergence time even in long-term diverse allopatric species. Although Coyne and Orr (1989; 1997) proposed that pre-zygotic isolation evolves more rapidly than post-zygotic isolation when allopatric taxa become sympatric, we found no evidence that pre-pollination isolation increased with divergence time in exotic-native Sagittaria species pairs.
We found clear molecular evidence for experimental hybrids between Sagittaria platyphylla and two native species (S. trifolia and S. pygmaea), which may threaten genetic integrity of native species due to introgression and potential genetic swamping (Ayres et al., 1999; Kellner et al., 2012; Ma et al., 2019). Indeed, HP transfer and putative natural hybrids with intermediate morphological characters between parental species was detected in three sympatric populations of S. platyphylla and S. trifolia (Zou Ting–Ting, unpublished data). In addition to hybridization risk, the excellent clonal propagation of S. platyphylla may cause interspecific competition for resources (Zhang et al., 2007; Sánchez-Vilas et al., 2011). Therefore, we highlight the need for possible conservation strategies to remove S. platyphylla and putative hybrids that are exposed to native species (especially endemic and endangered species) on a relatively large spatial scale. Furthermore, although S. montevidensis was incapable of hybridizing with native congeners, its larger floral display and higher pollen production indicates that the possibility for reduced reproductive success in native congeners via pollinator competition (Dietzsch et al., 2011; Beans and Roach, 2015) or ovule consumption (interspecific seed discounting) (Burgess et al., 2008; Nishida et al., 2014) exists. Further studies should focus on such potential reproductive interference from alien species to native species, as proposed by Moreira-Hernández and Muchhala (2019).
5. ConclusionWe found that multiple barriers act in concert to maintain species boundaries between alien and native Sagittaria species. Our results revealed that post-pollination barriers contributed more to total RI in native species, while pre-pollination barriers played a stronger role in total RI for two introduced species. Our results provide a novel perspective for comprehending different barriers for secondary contact species. S. platyphylla had strong hybridization proneness with native S. trifolia and S. pygmaea, which may pose a significant threat. By contrast, even though S. montevidensis was unlikely to be cross-compatible with all alien species, it still had the potential to interfere with native species' reproductive success. Thus, we recommend detecting natural hybrids and drafting an effective conservation action plan to remove the invasive species within the distribution range of native species.
AcknowledgementsWe thank Xiaolei Yu for helping with the sequential analysis. We thank Xin Zan for the help in the field. We are grateful to Chun-hui Wang for her valuable advice. This study was supported by the National Science Foundation of China (31970250).
Author contributions
T.-T. Zou and X.-F. Wang conceived and planned the study. T.-T. Zou, S.-T. Lyu, Q.-L. Jiang, and S.-H. Shang conducted the experiments. T.-T. Zou and S.-T. Lyu conducted statistical analyses and wrote the manuscript.
Declaration of competing interest
This manuscript has not been published or presented elsewhere in part or in entirety and is not under consideration by another journal. We have read and understood your journal's policies, and we believe that neither the manuscript nor the study violates any of these. There are no conflicts of interest to declare.
Appendice A and B. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.pld.2022.10.001.
Adair, R., Keener, B., Kwong, R., et al., 2012. The biology of Australian weeds 58. Sagittaria platyphylla (Engelmann) J.G.Smith, S. montevidensis Cham. & Schltdl. and S. calycina Engelmann. Plant Protect. Q., 27: 47-58. |
Ainouche, M.L., Fortune, P.M., Salmon, A., et al., 1999. Hybridization, polyploidy and invasion: lessons from Spartina (Poaceae). Biol. Invasions, 11: 1159-1173. |
Archibald, J.K., Mort, M.E., Crawford, D.J., et al., 2005. Life history affects the evolution of reproductive isolation among species of Coreopsis (Asteraceae). Evolution, 59: 2362-2369. |
Ayres, D.R., Garcia-Rossi, D., Davis, H.G., et al., 1999. Extent and degree of hybridization between exotic (Spartina alterniflora) and native (S. foliosa) cordgrass (Poaceae) in California, USA determined by random amplified polymorphic DNA (RAPDs). Mol. Ecol., 8: 1179-1186. |
Baack, E., Melo, M.C., Rieseberg, L.H., et al., 2015. The origins of reproductive isolation in plants. New Phytol., 207: 968-984. DOI:10.1111/nph.13424 |
Baldwin, B.G., 1992. Phylogenetic utility of the internal transcribed spacers of nuclear ribosomal DNA in plants: an example from the Compositae
. Mol. Phylogenet. Evol., 1: 3-16. |
Bandelt, H.-J., Forster, P., Röhl, A., 1999. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol., 16: 37-48. DOI:10.1093/oxfordjournals.molbev.a026036 |
Beans, C.M., Roach, D.A., 2015. An invasive plant alters pollinator-mediated phenotypic selection on a native congener. Am. J. Bot., 102: 50-57. DOI:10.3732/ajb.1400385 |
Bertin, R.I., 1990. Paternal success following mixed pollinations of Campsis radicans
. Am. Midl. Nat., 124: 153-163. DOI:10.2307/2426088 |
Brock, M.T., 2009. Prezygotic barriers to gene flow between Taraxacum ceratophorum and the invasive Taraxacum officinale (Asteraceae). Oecologia, 161: 241-251. DOI:10.1007/s00442-009-1383-0 |
Brys, R., Broeck, A.V., Mergeay, J., et al., 2014. The contribution of mating system variation to reproductive isolation in two closely related Centaurium species (Gentianaceae) with a generalized flower morphology. Evolution, 68: 1281-1293. DOI:10.1111/evo.12345 |
Brys, R., van Cauwenberghe, J., Jacquemyn, H., et al., 2016. The importance of autonomous selfing in preventing hybridization in three closely related plant species. J. Ecol., 104: 601-610. DOI:10.1111/1365-2745.12524 |
Burgess, K.S., Morgan, M., Husband, B.C., 2008. Interspecific seed discounting and the fertility cost of hybridization in an endangered species. New Phytol., 177: 276-284. DOI:10.1111/j.1469-8137.2007.02244.x |
Carney, S.E., Hodges, S.A., Arnold, M.L., 1996. Effects of differential pollen-tube growth on hybridization in the Louisiana irises. Evolution, 50: 1871-1878. |
Castro, M., Loureiro, J., Husband, B.C., et al., 2020. The role of multiple reproductive barriers: strong post-pollination interactions govern cytotype isolation in a tetraploid-octoploid contact zone. Ann. Bot., 126: 991-1003. DOI:10.1093/aob/mcaa084 |
Chen, J-K., 1989. Systematic and Evolutionary Botanical Studies on Chinese Sagittaria. Wuhan: Wuhan University Press.
|
Coyne, J.A., Orr, H.A., 1989. Patterns of speciation in Drosophila
. Evolution, 43: 362-381. |
Coyne, J.A., Orr, H.A., 1997. "Patterns of speciation in Drosophila" revisited. Evolution, 51: 295-303. |
Dell'olivo, A., Hoballah, M.E., Gubitz, T., et al., 2011. Isolation barriers between Petunia axillaris and Petunia integrifolia (Solanaceae). Evolution, 65: 1979-1991. DOI:10.1111/j.1558-5646.2011.01279.x |
Diaz, A., Macnair, M.R., 1999. Pollen tube competition as a mechanism of prezygotic reproductive isolation between Mimulus nasutus and its presumed progenitor M. guttatus. New Phytol., 144: 471-478. DOI:10.1046/j.1469-8137.1999.00543.x |
Dietzsch, A.C., Stanley, D.A., Stout, J.C., 2011. Relative abundance of an invasive alien plant affects native pollination processes. Oecologia, 167: 469-479. DOI:10.1007/s00442-011-1987-z |
Ellstrand, N.C., Schierenbeck, K.A., 2006. Hybridization as a stimulus for the evolution of invasiveness in plants?. Euphytica, 148: 35-46. DOI:10.1007/s10681-006-5939-3 |
Huang, S.-Q., 2003. Flower dimorphism and the maintenance of andromonoecy in Sagittaria guyanensis ssp. lappula (Alismataceae). New Phytol., 157: 357-364. |
Huang, S.-Q., Shi, X.-Q., 2013. Floral isolation in Pedicularis: how do congeners with shared pollinators minimize reproductive interference?. New Phytol., 199: 858-865. DOI:10.1111/nph.12327 |
Huang, S.-Q., Song, N., Wang, Q., et al., 2000. Sex expression and the evolutionary advantages of male flowers in an andromonoecious species, Sagittaria guyanensis subsp. lappula (Alismataceae). Acta Bot. Sin., 42: 1108-1114. |
Huang, S.-Q., Tang, L.-L., Sun, J.-F., et al., 2006. Pollinator response to female and male floral display in a monoecious species and its implications for the evolution of floral dimorphism. New Phytol., 171: 417-424. DOI:10.1111/j.1469-8137.2006.01766.x |
Jewell, C., Papineau, A.D., Freyre, R., et al., 2012. Patterns of reproductive isolation in Nolana (Chilean bellflower). Evolution, 66: 2628-2636. DOI:10.1111/j.1558-5646.2012.01607.x |
Keller, B., Ganz, R., Mora-Carrera, E., et al., 2021. Asymmetries of reproductive isolation are reflected in directionalities of hybridization: integrative evidence on the complexity of species boundaries. New Phytol., 229: 1795-1809. DOI:10.1111/nph.16849 |
Kellner, A., Ritz, C.M., Wissemann, V., 2012. Hybridization with invasive Rosa rugosa threatens the genetic integrity of native Rosa mollis
. Bot. J. Linn. Soc., 170: 472-484. DOI:10.1111/j.1095-8339.2012.01298.x |
Knope, M.L., Pender, R.J., Crawford, D.J., et al., 2013. Invasive congeners are unlikely to hybridize with native Hawaiian Bidens (Asteraceae). Am. J. Bot., 100: 1221-1226. DOI:10.3732/ajb.1300018 |
Larcombe, M.J., Silva, J.C., Tilyard, P., et al., 2016. On the persistence of reproductive barriers in Eucalyptus: the bridging of mechanical barriers to zygote formation by F1 hybrids is counteracted by intrinsic post-zygotic incompatibilities. Ann. Bot., 118: 431-444. DOI:10.1093/aob/mcw115 |
Levin, D.A., Francisco-Ortega, J., Jansen, R.K., 1996. Hybridization and the extinction of rare plant species. Conserv. Biol., 10: 10-16. |
Librado, P., Rozas, J., 2009. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics, 25: 1451-1452. DOI:10.1093/bioinformatics/btp187 |
Lowry, D.B., Modliszewski, J.L., Wright, K.M., et al., 2008. The strength and genetic basis of reproductive isolating barriers in flowering plants. Philos. Trans. R. Soc. B-Biol. Sci., 363: 3009-3021. DOI:10.1098/rstb.2008.0064 |
Lyu, N., Du, W., Wang, X.-F., 2016. Unique growth paths of heterospecific pollen tubes result in late entry into ovules in the gynoecium of Sagittaria (Alismataceae). Plant Biol., 19: 108-114. |
Ma, Y.-P., Xie, W.-J., Sun, W.-B., et al., 2016. Strong reproductive isolation despite occasional hybridization between a widely distributed and a narrow endemic Rhododendron species. Sci. Rep., 6: 19146. |
Ma, Y.P., Marczewski, T., Xue, D., et al., 2019. Conservation implications of asymmetric introgression and reproductive barriers in a rare primrose species. BMC Plant Biol., 19: 286. |
Mack, R.N., Simberloff, D., Mark Lonsdale, W., et al., 2000. Biotic Invasions: causes, epidemiology, global consequences, and control. Ecol. Appl., 10: 689-710. |
Martin, F.D., 1970. Pollen germination on foreign stigmas. Bull. Torrey Bot. Club, 97: 1-6. DOI:10.2307/2483984 |
Matallana, G., Oliveira, P.E., da Silva, P.R.R., et al., 2016. Post-pollination barriers in an assemblage of Bromeliaceae in south-eastern Brazil. Bot. J. Linn. Soc., 181: 521-531. DOI:10.1111/boj.12406 |
Matsumoto, T.K., Hirobe, M., Sueyoshi, M., et al., 2021. Selective pollination by fungus gnats potentially functions as an alternative reproductive isolation among five Arisaema species. Ann. Bot., 127: 633-644. DOI:10.1093/aob/mcaa204 |
McFarlane, S.E., Pemberton, J.M., 2019. Detecting the true extent of introgression during anthropogenic hybridization. Trends Ecol. Evol., 34: 315-326. |
Miguez, M.B., Amela García, M.T., 2019. Breeding system, floral biology and pollinators of Sagittaria montevidensis subsp. montevidensis Cham. et Schlech. (Alismataceae). Aquat. Bot., 158. |
Minnaar, C., de Jager, M.L., Anderson, B., 2019. Intraspecific divergence in floral-tube length promotes asymmetric pollen movement and reproductive isolation. New Phytol., 224: 1160-1170. DOI:10.1111/nph.15971 |
Montgomery, B.R., Soper, D.M., Delph, L.F., 2010. Asymmetrical conspecific seed-siring advantage between Silene latifolia and S. dioica. Ann. Bot., 105: 595-605. DOI:10.1093/aob/mcq013 |
Morales, C.L., Traveset, A., 2008. Interspecific pollen transfer: magnitude, prevalence and consequences for plant Fitness. Crit. Rev. Plant Sci., 27: 221-238. DOI:10.1080/07352680802205631 |
Moreira-Hernández, J.I., Muchhala, N., 2019. Importance of pollinator-mediated interspecific pollen transfer for angiosperm evolution. Annu. Rev. Ecol. Evol. Syst., 50: 191-217. DOI:10.1146/annurev-ecolsys-110218-024804 |
Munguía-Rosas, M.A., Jácome-Flores, M.E., 2020. Reproductive isolation between wild and domesticated chaya (Cnidoscolus aconitifolius) in sympatry. Plant Biol., 22: 932-938. DOI:10.1111/plb.13140 |
Ndlovu, M.S., Coetzee, J.A., Nxumalo, M.M., et al., 2020. The establishment and rapid spread of Sagittaria platyphylla in South Africa. Water, 12: 1472. DOI:10.3390/w12051472 |
Nishida, S., Kanaoka, M.M., Hashimoto, K., et al., 2014. Pollen-pistil interactions in reproductive interference: comparisons of heterospecific pollen tube growth from alien species between two native Taraxacum species. Funct. Ecol., 28: 450-457. DOI:10.1111/1365-2435.12165 |
Owens, G.L., Rieseberg, L.H., 2014. Hybrid incompatibility is acquired faster in annual than in perennial species of sunflower and tarweed. Evolution, 68: 893-900. DOI:10.1111/evo.12297 |
Paudel, B.R., Burd, M., Shrestha, M., et al., 2018. Reproductive isolation in alpine gingers: how do coexisting Roscoea (R. purpurea and R. tumjensis) conserve species integrity?. Evolution, 72: 1840-1850. DOI:10.1111/evo.13546 |
Qiu, J., Gao, C., Wei, H.L., et al., 2021. Flowering biology of Rhododendron pulchrum
. Horticulturae, 7: 508. DOI:10.3390/horticulturae7110508 |
Ramsey, J., Bradshaw, H.D., Schemske, D.W., 2003. Components of reproductive isolation between the monkeyflowers Mimulus lewisii and M. cardinalis (Phrymaceae). Evolution, 57: 1520-1534. |
Rozas, J., Ferrer-Mata, A., Sánchez-DelBarrio, J.C., et al., 2017. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol., 34: 3299-3302. DOI:10.1093/molbev/msx248 |
Sánchez-Vilas, J., Turner, A., Pannell, J.R., 2011. Sexual dimorphism in intra- and interspecific competitive ability of the dioecious herb Mercurialis annua
. Plant Biol., 13: 218-222. DOI:10.1111/j.1438-8677.2010.00408.x |
Sobel, J.M., Chen, G.F., 2014. Unification of methods for estimating the strength of reproductive isolation. Evolution, 68: 1511-1522. DOI:10.1111/evo.12362 |
Tamura, K., Stecher, G., Kumar, S., 2021. MEGA11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol., 38: 3022-3027. DOI:10.1093/molbev/msab120 |
Tang, J., Sun, S.-G., Haung, S.-Q., 2022. Experimental sympatry suggests geographic isolation as an essential reproductive barrier between two sister species of Pedicularis
. J. Syst. Evol.: 1-12. |
Todesco, M., Pascual, M.A., Owens, G.L., et al., 2016. Hybridization and extinction. Evol. Appl., 9: 892-908. DOI:10.1111/eva.12367 |
Tong, Z.-Y., Huang, S.-Q., 2016. Pre- and post-pollination interaction between six co-flowering Pedicularis species via heterospecific pollen transfer. New Phytol., 211: 1452-1461. DOI:10.1111/nph.14005 |
Walsh, P.S., Metzger, D.A., Higushi, R., 1991. Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques, 10: 506-513. |
Wang, H., Wang, Q., Bowler, P., et al., 2016. Invasive aquatic plants in China. Aquat. Invasions, 11: 1-9. |
Wang, H., Xiao, K.Y., Wu, Z.G., et al., 2020. Delta arrowhead (Sagittaria platyphylla) in the Yangtze River: an invasive aquatic plant and the potential ecological consequences. BioInvasions Rec., 9: 618-626. DOI:10.3391/bir.2020.9.3.17 |
Wang, Q.F., Li, W., Wang, G.X., et al., 2021. Aquatic Plant of China. Wuhan: Hubei Science and Technology Press.
|
Wang, X.-F., Tan, Y.-Y., Chen, J.-H., et al., 2006. Pollen tube reallocation in two preanthesis cleistogamous species, Ranalisma rostratum and Sagittaria guyanensis ssp. lappula (Alismataceae). Aquat. Bot., 85: 233-240. |
Zaya, D.N., Leicht-Young, S.A., Pavlovic, N.B., et al., 2015. Genetic characterization of hybridization between native and invasive bittersweet vines (Celastrus spp.). Biol. Invasions, 17: 2975-2988. DOI:10.1007/s10530-015-0926-z |
Zhang, Z.H., Rengel, Z., Meney, K., 2007. Growth and resource allocation of Canna indica and Schoenoplectus validus as affected by interspecific competition and nutrient availability. Hydrobiologia, 589: 235-248. |
Zhong, S., Liu, M.L., Wang, Z.J., et al., 2019. Cysteine-rich peptides promote interspecific genetic isolation in Arabidopsis
. Science, 364: 851. |
Zou, T.-T., 2022. ITS sequence of native and alien Sagittaria species and their hybrids. Figshare. Dataset. |
Zou, T.-T., Wang, C.-H., Lyu, S.-T., et al., 2022. Effects of heterospecific pollen on stigma behavior in Campsis radicans: causes and consequences. Am. J. Bot., 109: 1-12. |