



b. Key Laboratory of Tropical Forest Ecology,Xishuangbanna Tropical Botanical Garden,Chinese Academy of Sciences,Mengla 666303,China;
c. Senckenberg Research Institute/BiK F (Loewe),Senckenberganlage 25,60325 Frankfurt am Main,Germany;
d. State Key Laboratory of Paleobiology and Stratigraphy,Nanjing Institute of Geology and Paleontology,Chinese Academy of Sciences,Nanjing 210008,China;
e. Graduate University of the Chinese Academy of Sciences,Beijing 100049,China;
f. Steinmann Institute,University of Bonn,Nussallee 8,53115 Bonn,Germany
Yunnan in southwestern China has long been recognized as a hotspot for plant diversity(Wu, 1988; López-Pujol et al., 2006, 2011; Ruth et al., 2008; Turkington and Harrower, 2016). It is home to approximately 16, 000 higher plant species grouped into more than 2300 genera within 244 families, representing 6% of the world's total diversity of higher plant species(Wu, 1979), even though the region accounts for only 4% of the Chinese land surface. Nearly half of these plant species are endemic to Yunnan. In addition, almost all ecosystem types within China, including tropical seasonal rain forest, evergreen broad-leaved forest, sclerophyllous evergreen broad-leaved forest, deciduous broad-leaved forest, both warm and cold temperate coniferous forests, dry and warm valley scrubs as well as subalpine and alpine scrubs can be found in Yunnan(Writing Group of Yunnan Vegetation, 1987). This high diversity of plant taxa and vegetation types qualifies Yunnan as an outstanding place for botanical research. Yunnan also exhibited exceptional botanical richness in the geological past as revealed by the rich fossil record. To date, around 20 megafossil floras are known from the Cenozoic of Yunnan, and hundreds of fossil species, covering ferns, gymnosperms and angiosperms, have been documented. A summary of past plant diversity based on palaeobotanical data affords a better understanding of the modern botanical richness in this region.
In the present review, we compile palaeobotanical data from literature to document plant diversity in Yunnan throughout the Cenozoic. Only fossil taxa represented by megafossils are considered, i.e., leaves, wood, fruits, seeds and flowers. Pollen records are not considered, because they can be transported to Yunnan from elsewhere and cannot be identified to a fine enough taxonomic resolution. We first briefly introduce the research history of palaeobotany in Yunnan, then present known fossil families, genera and species to depict the diversity of Cenozoic plants and ecosystems, and analyze the inferred floristic characters and changes. We also summarize the Neogene climate evolution in Yunnan and related extinction events exemplified for selected plant taxa. The present review aims to facilitate access to the rich palaeobotanical record of Yunnan for both palaeobotanists and botanists.
2. Research history of palaeobotany in YunnanAs a montane region, Yunnan has more than 150 Cenozoic sedimentary basins, many of which have yielded Cenozoic strata that bear plant fossils(Ge and Li, 1999). The region archives at least 20 megafossil floras, ranging from the early Oligocene to late Pliocene(Fig. 1). However, these fossil floras rarely attracted attention before the beginning of the last century. In 1920, Colani reported the first plant fossils recovered from Miocene sediments along the former railway connecting Kunming and Hanoi(Colani, 1920), but interest was transient as no more palaeobotanical studies followed for the next 57 years. In 1978, several Chinese paleobotanists, including Tao Jun-Rong, Guo Shuang-Xing and Li Hao-Ming, jointly published a monograph entitled “Cenozoic Plants of China”(Writing Group of the Cenozoic Plants of China, 1978). In that monograph, 301 plant fossil species, covering mosses, ferns, gymnosperms and angiosperms, were described systematically. Among them, 76 species came from five floras in Yunnan: the Oligocene Jinggu flora, the Miocene Duotan flora, the early Miocene Shuanghe flora, the late Miocene Xiaolongtan flora, and the late Pliocene Eryuan flora. In the 1980s, Tao and Chen(1983)described the late Miocene Bangmai flora, also known as the Lincang flora, from southwestern Yunnan which comprised 59 fossil species grouped into 48 genera within 28 families; Tao(1986)reported the late Pliocene Fudong flora from northwestern Yunnan which yielded 17 species grouped into four genera of three families. Unfortunately, published mostly in Chinese, these works have had limited international recognition.

|
| Fig. 1 Map showing the distribution of Cenozoic fossil floras in Yunnan, southwestern China |
During the last two decades, 10 Cenozoic megafossil floras from Yunnan have been studied as doctoral or master's research projects, e.g., the late Miocene Xianfeng flora, the late Pliocene Nanbanbang, Fudong, Longmen and Tuantian floras. A few are still being investigated for the subject of doctoral research, e.g., the middle Miocene Wenshan and Maguan floras(formerly dated as late Miocene, but recently moved to middle Miocene according to lake geochemical evidence; Lebreton-Anberrée et al., 2016). To the best of our knowledge, at least eight doctoral and two master's theses are freely accessible at universities or institutions in China, e.g., Lanzhou University, Kunming Institute of Botany, Chinese Academy of Sciences(Zhao, 2002; Xie, 2007; Wu, 2010; Su, 2010; Xing, 2010; Huang, 2012; Hu, 2013; Li, 2015; Wang, 2016; Zhu, 2016). Such theses usually present a full list of identified plant taxa that more or less reflects a given fossil flora(Wu, 2010; Huang, 2012; Zhu, 2016). Some of the authors have also interpreted the palaeovegetation or palaeoclimate accordingly(Wu, 2010; Su, 2010; Xing, 2010). Based on detailed palaeobotanical research, hundreds of fossil species have been published in a variety of international journals(e.g., Zhao et al., 2004; Xie et al., 2010; Xing et al., 2010, 2013; Su et al., 2011, 2013b; Huang et al., 2012, 2013; Wu et al., 2012; Hu et al., 2014; Jia et al., 2015; Li et al., 2015; Zhu et al., 2015, 2016; Huang et al., 2016a; Fig. 2). In addition to those targeted within doctoral or master's projects, research has begun on other fossil floras, such as the Oligocene Jinggu flora, the middle Miocene Sanzhangtian flora, and the late Miocene Shuitangba flora, and tens of fossil species have been described(e.g., Wen et al., 2013; Jacques et al., 2013; Wang et al., 2013; Meng et al., 2014, 2015; Xie et al., 2014; Zhang et al., 2015a, 2015b; Liang et al., 2016; Su et al., 2016; Fig. 2). It seems that the recent decades have witnessed an upward surge in palaeobotanical research in Yunnan.

|
| Fig. 2 Selected fossil taxa comprising leaves, wood, fruits and seeds from the Cenozoic of Yunnan. Scale bars ¼ 10 mm for images 1e3, 5 and 8e10; 0.5 mm for images 4, 7, 11 and 12.1. Drynaria callispora(Polypodiaceae)from the late Pliocene Longmen flora(Su et al., 2011). 2. Calocedrus shengxianensis(Cupressaceae)from the middle Miocene Wenshan flora(Zhang et al., 2015b). 3. Tsuga xianfengensis(Pinaceae)from the late Miocene Xianfeng flora(Xing et al., 2013). 4. Fossil wood of Pinus uniseriata(Pinaceae)from the Xianfeng flora(Wang et al., in press). 5. Mahonia mioasiatica(Berberidaceae)from the Wenshan flora(Huang et al., 2016a). 6. Quercus tenuipilosa(Fagaceae)from the late Pliocene Hunshuitang flora(Hu et al., 2014). 7. A fossil seed of Ranunculus(Ranunculaceae)from the Nanbanbang flora(unpublished data). 8. Rosa fortuita(Rosaceae)from the Wenshan flora(Su et al., 2016). 9. Celastrus caducidentatus(Celastraceae)from the middle Miocene Sanzhangtian flora(Liang et al., 2016). 10. Bauhinia wenshanensis(Fabaceae)from the Wenshan flora(Meng et al., 2014). 11. Eurya stigmosa(Theaceae)from the late Pliocene Nanbanbang flora(Zhu et al., 2016). 12. Aralia stratosa(Araliaceae)from the late Pliocene Fudong flora(Zhu et al., 2015). |
Up to now, approximately 386 plant species(including morphospecies)belonging to 170 genera within 66 families have been documented from the Cenozoic of Yunnan and most of them are from the Neogene(see the Appendix). They include ferns, gymnosperms and angiosperms, of which ferns and gymnosperms are relatively few.
Fossil ferns from Yunnan comprise only nine species placed in five genera of four families: Davalliaceae, Equisetaceae, Osmundaceae and Polypodiaceae. The genus Drynaria in Polypodiaceae is the most diverse and has four different fossil species, which all came from the Neogene of western Yunnan(Su et al., 2011; Wu et al., 2012; Wen et al., 2013; Huang et al., 2016b). This may from a historical view explicate the establishment of the modern Drynaria diversity in the Hengduan Mountains, western Yunnan(Huang et al., 2016b). Fossil gymnosperms from Yunnan fall into 24 species grouped into 10 genera of two families, Pinaceae and Cupressaceae. The former has 15 fossil species belonging to four genera(Cedrus, Picea, Pinus and Tsuga), while the latter has nine fossil species belonging to six genera(Calocedrus, Cephalotaxus, Cupressus, Platycladus, Sequoia and Taiwania).
Similar to modern Yunnan, angiosperms were the most diverse plant group throughout the Cenozoic. A total of 353 fossil species of angiosperms grouped into 155 genera within 60 families are known from megafossil remains. At the family level, Fabaceae has 23 genera with fossil records and represents the most genus-rich family in the fossils, followed by Lauraceae which has 11 fossil genera(Table 1). Other angiosperm families having at least five genera of fossil plants are Rosaceae(8 genera), Fagaceae(7 genera), Juglandaceae(6 genera), Malvaceae(6 genera)and Rutaceae(5 genera). Angiosperm families having four genera of fossil plants include Anacardiaceae, Betulaceae, Hamamelidaceae, Poaceae and Rhamnaceae. Most families are represented by only one genus with fossil evidence, e.g., Buxaceae, Caryophyllaceae, Elaeagnaceae and Moraceae(Table 1). Further fossil finds will no doubt reveal more. For their diversity at the species level, Fagaceae having 79 fossil species is the most species-rich family, followed by Fabaceae and Lauraceae with 38 and 33 fossil species, respectively. There are seven other families having 10 or more fossil species, namely Juglandaceae(14 species), Ulmaceae(13 species), Sapindaceae(13 species), Salicaceae(12 species), Betulaceae(11 species), Anarcardiaceae(11 species)and Rosaceae(10 species). Most families, however, have only one or two fossil species, e.g., Euphorbiaceae, Oleaceae, Nymphaeaceae and Celastraceae(Table 1). Based on the above summary, Fabaceae, Fagaceae, Lauraceae, Juglandaceae and Rosaceae were among the most diversified families in the Cenozoic of Yunnan either at the generic level or at the species level. Today, Fagaceae and Lauraceae are two of the major families contributing to evergreen broad-leaved forests in Yunnan as well as in other parts of southern China(Writing Group of Yunnan Vegetation, 1987), and Fabaceae is among the most common families in tropical and subtropical forests in Yunnan. Rich fossil records of Fabaceae, Fagaceae and Lauraceae suggest that their dominance in the present evergreen broad-leaved forests of Yunnan seems to have been largely established at least by the Neogene. In addition, many other families that are widely seen in the modern broad-leaved forests were also found as fossils, e.g., Juglandaceae, Ulmaceae, Sapindaceae, Salicaceae, Betulaceae and Rosaceae. This suggests that themodern floristic composition at the family level in Yunnan was probably formed in the Neogene or before. Interestingly, some families such as Magnoliaceae and Theaceae that are important and frequent components today are not well represented in the fossil record(Table 1).
With respect to the species diversity of genera, six angiosperm genera have more than 10 fossil species, i.e., Quercus, Castanopsis, Lithocarpus, Acer, Cyclobalanopsis and Cinnamomum(Table 2; Fig. 3). Among them, Quercus is obviously the richest and has 36 fossil species; Castanopsis, Lithocarpus and Acer have 14, 13 and 11 fossil species, respectively; and Cyclobalanopsis and Cinnamomum each has 10 fossil species. It is worth explaining that different classifications of Quercus and Cyclobalanopsis exist. Some botanists consider Quercus and Cyclobalanopsis as two separate genera distinguished by the imbricate-scaled or lamellate structure of the e.g., Betula and Ficus both with four fossil species, Alnus and Carpinus with three, and Lindera and Mahonia with two(Table 2). Most of the genera have only one fossil species, e.g., Buxus, Citrus, Cornus and Sambucus. Notably, the genera that were species diverse in the Cenozoic mainly belong to Fagaceae(Querucs, Castanopsis, Lithocarpus and Cyclobalanopsis)and Lauraceae(Cinnamomum, Phoebe and Machilus), and as already mentioned, they are still diverse in modern forests of Yunnan. Other genera that have good fossil records are also important components of modern broad-leaved forests in Yunnan, e.g., Acer, Ulmus, Myrica, Rhus and Salix. This therefore suggests the idea that the composition of Neogene evergreen broad-leaved forests in this regionwas already similar to that today at the generic level.

|
| Fig. 3 Species diversity of the fossil-richest genera from the Cenozoic of Yunnan |
Molecular studies have been repeatedly interpreted as meaning that numerous plant groups in Yunnan experienced species diversification as a result of increasing environmental complexity(e.g., Wang et al., 2005; Yu et al., 2015). The other side of the historical emergence of new lineages, however, is species extinction. Although most families and genera recorded as fossils still exist, some genera disappeared from Yunnan after the Neogene. To date, there are seven genera with fossil evidence we do not encounter in the natural vegetation of the region. They are Cedrus, Sequoia, Berryophyllum, Cedrelospermum, Palaeocarya, Podocarpium and Wataria(Fig. 4).

|
| Fig. 4 Selected fossil taxa now extinct in Yunnan. Scale bars ¼ 10 mm for all images. 1. Seed scale of Cedrus angusta(Pinaceae)from the late Pliocene Longmen flora(Su et al., 2013b). 2, 3. Winged fruits of Cedrelospermum asiaticum(Ulmaceae)from the middle Miocene Maguan flora(Jia et al., 2015). 4. Leafy shoot of Sequoia maguanensis(Cupressaceae; Zhang et al., 2015a)from the middle Miocene Wenshan flora. |
Cedrus is a small genus of four extant species restricted to Mediterranean regions and the western Himalayas(Maheshwari and Biswas, 1970). In modern Yunnan, no Cedrus grows as a natural population. However, a recent fossil discovery of seeds and cone scales indicate that Cedrus occurred in northwestern Yunnan during the late Pliocene(Su et al., 2013b). The genus must have disappeared from this region soon after(Su et al., 2013b), as is also evidenced by the decline in its pollen occurrences(Tong et al., 1990; Wang and Shu, 2004; Xiao et al., 2010). Sequoia is a genus now endemic only to the western coast of North America(Waston and Eckenwalder, 1993). Like the situation with Cedrus, Sequoia also has a fossil representative in Yunnan. Fossils of cones and leafy shoots discovered from the middle Miocene Maguan flora in southeastern Yunnan closely resemble modern Sequoia, indicating its occurrence in the region at that time(Zhang et al., 2015a). The genus probably disappeared from Yunnan after the middle Miocene. This raises the question as to why Cedrus and Sequoia went extinct in Yunnan but survive in other regions. Investigations by Su et al.(2013b)and Zhang et al.(2015a)have demonstrated that the aridification of the dry season, associated with monsoon intensification, might have played a major role. Today, Yunnan has dry conditions from late autumn to early spring, whereas Mediterranean regions and the western coast of North America both have wet winters and springs. As seeds of Cedrus and Sequoia are recalcitrant, they germinate immediately upon their maturity in the autumn(Olson et al., 1990; Kumar et al., 2011). Unfortunately, dry conditions after autumn may prevent their seeds from germinating or their seedlings from developing new leaves. In contrast to modern climates, in the Neogene Yunnan had relatively humid conditions in the dry season(Xia et al., 2009; Sun et al., 2011; Xing et al., 2012; Su et al., 2013a; Huang et al., 2015a). Humid and suitable habitats might explain the historical existence of the two genera in this region. As precipitation in the dry season decreased, likely linked to the Asian monsoon intensification, as is elaborated below, the dry winter and spring ultimately led to the regional extinction of species that require a moister dry season(Su et al., 2013b; Zhang et al., 2015a).
Unlike Cedrus and Sequoia that have no living relatives in Yunnan but survive in other regions, the remaining five genera only have fossil representatives; in other words, they have gone extinct globally. Berryophyllum is an extinct genus morphologically similar to extant Lithocarpus, Canstanopsis and Cyclobalanopsis of Fagaceae(Zhou, 1996). The genus was widely distributed in Europe and North America from the Paleocene to Miocene(Jones and Dilcher, 1988; Zhou, 1996). In Yunnan, it has been described only from the Oligocene Jinggu flora in southern Yunnan. Cedrelospermum(Ulmaceae)is another extinct genus that was well represented in Europe(Hably and Thiébaut, 2002; and references therein)and North America(Manchester, 1987, 1989; Manchester and Tiffney, 2001). In Yunnan, one fossil species described as Cedrelospermum asiaticum has been reported from the middle Miocene Maguan flora in southeastern Yunnan(Jia et al., 2015). This species represents the first record of Cedrelospermum in Asia(Jia et al., 2015). Palaeocarya is morphologically similar to two modern genera, namely Engelhardia and Oreomunnea of Juglandaceae(Manchester, 1999). It has an excellent fossil record in the Northern Hemisphere, extending from the Pliocene deeply into the Eocene(Tanai and Uemura, 1983; Jähnichen et al., 1984; Manchester, 1987; Guo and Zhang, 2002; Jin, 2009; Meng et al., 2015). In Yunnan, Palaeocarya has been reported from the middle Miocene Wenshan flora in southeastern Yunnan(Meng et al., 2015)and the late Pliocene Tuantian flora in southwestern Yunnan(Xie et al., 2010). Podocarpium of Fabaceae is one of the most common legumes recorded from the Eocene to Pliocene(Xu et al., 2015). In Yunnan, the genus has one fossil species named P. poocarpum from the late Miocene Xiaolongtan flora in southern Yunnan(Guo and Zhou, 1992). The extinct genus Wataria, established by Terada and Suzuki(1998), is anatomically similar to Reeveia of Malvaceae. In Yunnan, Wateria yunnanica has been described from the middle Miocene Shengli flora in southern Yunnan on the basis of fossil wood(Li et al., 2015). This wood species represents the first fossil occurrence of the genus in China(Li et al., 2015).
5. Floristic characters and changesOf the 20 megafossil floras from Yunnan considered here, those of late Miocene and late Pliocene age have been well studied and they are relatively abundant(Fig. 1). Hence these floras may readily reflect floristic characters of the two time slices in the region as well as their changes. Fossil floras of other stratigraphical phases are not considered in the present analyses, as they have been insufficiently studied from a formal systematic viewpoint.
There are four late Miocene floras known from Yunnan on the basis of which we look at the lateMiocene floristic characteristics: the Shuitangba, Xianfeng, Bangmai and Xiaolongtan floras. The late Miocene Shuitangba flora in northeastern Yunnan is dominated by subtropical representatives of Quercus as revealed by the pollen record(Chang et al., 2015), and Euryale in the aquatic ecosystem as reflected by recently recovered carpological remains(Huang et al., 2015b). The flora contains several temperate elements such as Corylus and Staphylea, and only a few pantropical ones such as Zanthoxylum. Thus the Shuitangba flora might represent a northern subtropical flora. The late Miocene Xianfeng and Xiaolongtan floras in eastern Yunnan are both dominated by the evergreen fagaceous elements such as Cyclobalanopsis, Lithocarpus and Quercus, as well as Lauraceae(Xia et al., 2009; Xing et al., 2012). In the Xiaolongtan flora, tropical and subtropical Fabaceae are relatively abundant(Guo and Zhou, 1992; Xia et al., 2009), but the percentage of tropical components(30-50%)is still belowthe minimumlevel(60%)that defines a tropical floristic region(Zhu, 2013). Therefore, the Xianfeng and Xiaolongtan floras represent two subtropical floras. By contrast, the late Miocene Bangmai flora in southwestern Yunnan contains more tropical genera, accounting for 60% of the fossil assemblage(Guo, 2011). Moreover, several genera are mainly or even merely distributed in tropical regions, such as Mucuna, Murraya, Neocinnamomum, Ormosia, Piper and Terminalia(Guo, 2011). According to the definition of a tropical floristic region by Zhu(2013), the late Miocene Bangmai flora would qualify as tropical. Thus we conclude that in the late Miocene three known floristic regions occurred in Yunnan: a northern subtropical floristic region in the northeast, a subtropical floristic region in the east, and a tropical floristic region in the southwest(Fig. 5A).

|
| Fig. 5 Floristic regions in the late Miocene (A) and late Pliocene (B) of Yunnan (grey areas: no data). I. Tropical floristic region. II. Subtropical floristic region. III. Northern subtropical floristic region. IV. Subalpine floristic region. |
For the late Pliocene, seven fossil floras are known from Yunnan, i.e., the Nanbanbang, Fudong, Eryuan, Longmen, Tuantian, Yuanmou and Hunshuitang floras. The first four floras from the southern Hengduan Mountains in northwestern Yunnan are fairly distinguished by the dominance of mountain oaks, Quercus sect. Heterobalanus, suggesting sclerophyllous evergreen broad-leaved forests during the late Pliocene. This is well supported by the fact that seven out of the 20 fossil species from the Fudong flora belong to Quercus sect. Heterobalanus(Huang, 2012), and 1275 out of around 1500 fossil specimens from the Longmen flora belong to this section(Su, 2010). Today, sclerophyllous evergreen broad-leaved forests dominated by Quercus sect. Heterobalanus are widely distributed in northwestern Yunnan(Writing Group of Yunnan Vegetation, 1987). This pattern might have been largely established by the late Pliocene. Distinct from these four floras, the contemporary Tuantian flora in southwestern Yunnan was shown to contain many tropical to subtropical elements, accounting for about 53% of the recognized fossil taxa(Wu, 2010). They include Castanposis, Cyclobalanopsis and Quercus of Fagaceae, and Cinnamomum, Lindera and Machilus of Lauraceae(Wu, 2010), which collectively characterize a subtropical evergreen broad-leaved forest. The late Pliocene Hunshuitang flora in eastern central Yunnan is dominated by Cyclobalanopsis(Hu, 2013; Hu et al., 2014), also suggesting a subtropical evergreen broad-leaved forest. Hence in the late Pliocene of Yunnan, at least two kinds of floristic regions existed: sclerophyllous evergreen broad-leaved forests in the northwest that represented a subalpine floristic region, and subtropical evergreen broad-leaved forests in the southwest and eastern center that in each case represented a subtropical floristic region(Fig. 5B).
Based on the above discussions, Yunnan possibly witnessed different floristic features between the late Miocene and late Pliocene. This allows us to observe floristic changes from the late Miocene to late Pliocene through to today. In northern Yunnan, the proportion of temperate elements at the generic level has increased since the late Miocene(Jacques et al., 2014). This is best exemplified by the fact that sclerophyllous evergreen broad-leaved forests were apparently absent during the late Miocene, but were well established in the late Pliocene and are still seen today. In southern Yunnan, the percentage of tropical elements declined significantly, as was previously noticed by Jacques et al.(2014). Today, Yunnan is covered mainly by subtropical evergreen broad-leaved forests, with tropical forests being restricted to its southern most parts. Overall, it appears that thermophylic floras have moved southward since the late Miocene, purportedly in response to elevation changes(Jacques et al., 2014).
6. Areal type analyses of seed plantsResearch into spatial distribution patterns of plants can help botanists to better understand the origin of a flora and its relationship with others. In China, this kind of study began in 1965(Wu, 1965). In 1991, Wu(1991)summarized the results of previous studies and proposed a term named “areal type” to reveal the distribution of a genus of seed plants. In that summary, 15 areal types of seed plants distributed in China were defined and described(Wu, 1991). They are 1. Cosmopolitan, 2. Pantropic, 3. Tropical Asia and Tropical America disjunct, 4. Old World Tropic, 5. Tropical Asia and Tropical Australasia, 6. Tropical Asia to Tropical Africa, 7. Tropical Asia(IndoeMalaysia), 8. North Temperate, 9. East Asia and North America disjunct, 10. Old World Temperate, 11. Temperate Asia, 12. Mediterranean, West Asia to Central Asia, 13. Central Asia, 14. East Asia, and 15. Endemic to China(Wu, 1991;Wu et al., 2003). In the present review, we apply the definitions by Wu(1991)to analyze the areal types of seed plant genera from the Cenozoic of Yunnan. To achieve the best spatial resolution, we focus only on the late Miocene and late Pliocene floras, because of their relative abundance in Yunnan(Tables 3 and 4).
|
| Areal type | Fudong | Eryuan | Longmen | Tuantian |
| 1 | / | / | / | / |
| 2 | / | Celtis | / | Dioscorea, Ilex |
| 3 | / | / | / | Cinnamomum, Ormosia |
| 4 | / | / | / | Mallotus, Syzygium |
| 5 | / | / | / | Evodia |
| 6 | / | / | Garcinia | |
| 7 | Cyclobalanopsis | / | / | Cyclea, Cyclobalanopsis, Exbucklandia, Heterosmilax, Machilus, Rhodoleia |
| 8 | Acer, Berberis, | Acer, Pinus, | Acer, Cupressus, | Betula, Carpinus, |
| Picea, Pinus, | Populus, | Pinus, Populus, | Castanea, Cornus, | |
| Populus, | Quercus, | Quercus, | Fraxinus, Juglans, | |
| Quercus, Salix | Viburnum | Rhododendron, Salix | Myrica, Ulmus | |
| 9 | Desmodium, Lithocarpus | / | Lithocarpus | Berchemia, Castanopsis, Lindera, Mahonia, Robinia |
| 10 | / | Trapa | Trapa | / |
| 11 | / | / | / | / |
| 12 | / | / | / | / |
| 13 | / | / | / | / |
| 14 | / | / | / | / |
| 15 | / | / | / | / |
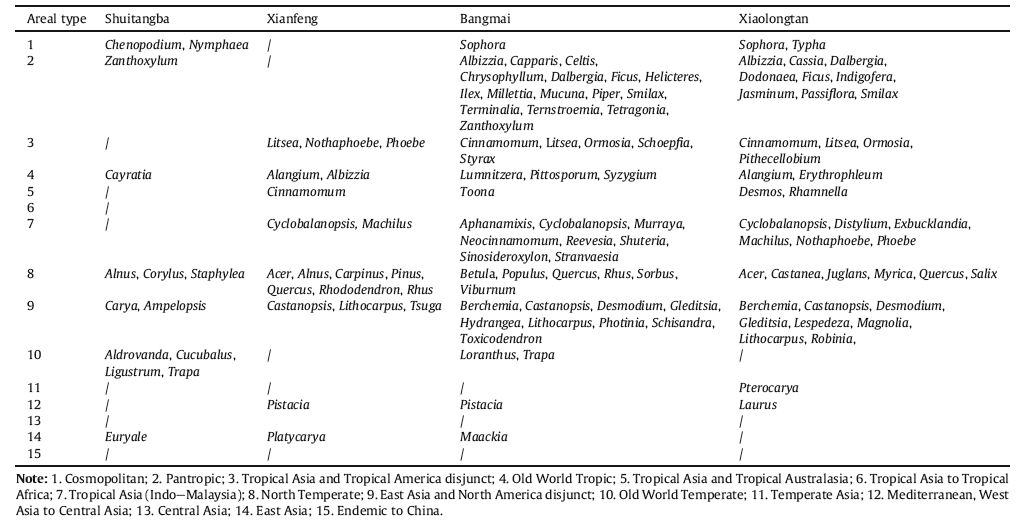
The late Miocene Shuitangba flora is represented by seven areal types at the generic level, the Old World Temperate type of which being the most abundant, followed by the North Temperate type. The late Miocene Xianfeng flora is dominated by the North Temperate type followed by the Tropical Asia and Tropical America disjunct type and the East Asia and North America disjunct type(Fig. 6). The late Miocene Bangmai and Xiaolongtan floras comprise several areal types with each including various genera. The Pantropic type frequently occurred in both floras, with the number of genera being 16 and 9, respectively. Other important areal types are the East Asia and North America disjunct type, the Tropical Asia(IndoeMalaysia)type and the North Temperate type. It is interesting that the Pantropic type and Tropical Asia(IndoeMalaysia)type are best to well represented in the Bangmai and Xiaolongtan floras in southern Yunnan, whereas they are rare to absent in the Shuitangba and Xianfeng floras in northern Yunnan(Fig. 6). Because these two areal types of seed plants reflect a tropical floristic composition, the Bangmai and Xiaolongtan floras may have a higher percentage of tropical elements than the Shuitangba and Xianfeng floras. Moreover, it should be noted that the Bangmai and Xiaolongtan floras have similar compositions of areal types. This seems to disagree with the suggested subdivision of floristic regions, namely that the late Miocene Bangmai flora represented a tropical floristic region while the late Miocene Xiaolongtan flora represented a subtropical floristic region. However, the Pantropic type of the former is considerably richer than the latter, which apparently supports our floristic region division. Hence it can be concluded that in the late Miocene southern Yunnan had far more Pantropic type elements than northern Yunnan.

|
| Fig. 6 Composition of areal types for four late Miocene floras from Yunnan |
To reconstruct the situation in the late Pliocene, four fossil floras can be used: the Fudong, Eryuan, Longmen and Tuantian floras. As is shown, the first three floras from northwestern Yunnan have only a few areal types, meaning low diversity of areal types(Fig. 7). Interestingly, they are all dominated by the North Temperate type, suggesting temperate floristic features. This is in line with results of our floristic analyses that northwestern Yunnan represented a subalpine floristic region. Different from the Fudong, Eryuan and Longmen floras, the Tuantian flora in southwestern Yunnan is much more diverse with regards to areal types(Fig. 7). Although the North Temperate type is also dominant, the Tropical Asia(IndoeMalaysia)type and the East Asia and North America disjunct type are frequent. Other areal types include the Pantropic type, the Tropical Asia and Tropical America disjunct type, the Old World Tropic type, the Tropical Asia and Tropical Australasia type, and the Tropical Asia to Tropical Africa type, which are found mainly in tropical zones. Therefore, the Tuantian flora is closer to a tropical flora than the remaining three, as is also indicated by our floristic analyses. In short, late Pliocene plant assemblages in northwestern Yunnan were dominated exclusively by the North Temperate type, while in southwestern Yunnan the Tropical Asia(IndoeMalaysia)type and the East Asia and North America disjunct type also played important roles.

|
| Fig. 7 Composition of areal types for four late Pliocene floras from Yunnan |
Because plants only exist in a climate zone that they can tolerate, they reflect climate characteristics of their distributional ranges. Although a fossil flora can tell us the general climate situation(e.g., the late Miocene Bangmai flora dominated by tropical elements indicates a warm climate), quantitative climate results allow a better understanding of past climate. As methodologies develop, fossil floras become valuable proxies for quantitative reconstructions of palaeoclimates by using appropriate approaches. In Yunnan, 10 megafossil floras have been investigated quantitatively regarding their palaeoclimates. They are exclusively Neogene in age, i.e., the early Miocene Shuanghe flora(Sun et al., 2011), the early to middle Miocene Mangdan flora(Zhao et al., 2004), the middle Miocene Tuantian flora(Sun et al., 2011), the late Miocene Xianfeng, Xiaolongtan and Bangmai floras(Xia et al., 2009; Jacques et al., 2011; Xing et al., 2012), and the late Pliocene Fudong, Longmen, Eryuan and Tuantian floras(Sun et al., 2011; Xie et al., 2012; Su et al., 2013a; Huang et al., 2015a). The reconstructed palaeoclimate results provide insights into climate changes and monsoon development in Yunnan throughout the Neogene, and thus help us to better understand the environmental conditions under which plant diversity evolved.
In the Miocene, estimated temperatures indicate that northern Yunnan was much warmer than it is now, suggesting a significant cooling trend to the present(Sun et al., 2011; Xing et al., 2012). For example, the mean annual temperature(MAT)of the early Miocene Shuanghe flora was estimated to be 13.8e21.7 ℃ by Coexistence Approach(CA)and 18.2 ± 3.6 ℃ by Leaf Margin Analysis(LMA), compared to 12.3 ℃ today(Sun et al., 2011). By contrast, temperatures in southern Yunnan have changed only moderately since the Miocene(Zhao et al., 2004; Xia et al., 2009). For example, the MAT estimated from the early to middle Miocene Mangdan flora using the CA was 18.8e20.5 ℃, close to 19-21 ℃ at present(Zhao et al., 2004). The observed different patterns of temperature changes between northern and southern Yunnan might be attributed to the difference in latitude or altitude or both. Northern Yunnan clearly had higher latitude than southern Yunnan, which supports a previous interpretation that the late Cenozoic cooling of the Northern Hemispherewas more pronounced in higher latitudes than in lower ones(Uhl et al., 2007; Utescher et al., 2011). On the other hand, northern Yunnan underwent significant surface uplift during the Neogene due to the southeastern extrusion of the Tibetan Plateau(Zhang and Ding, 2003; Clark et al., 2005), whereas southern Yunnan remained almost at the same level(Jacques et al., 2014). The considerable altitude increase probably was the other factor leading to the observed surface temperature decline in northern Yunnan, while the relatively stable elevation in southern Yunnan may explain the almost unchanged temperature level. The relevant importance of latitude versus altitude in controlling temperature changes, however, is currently unanswerable.
Apart from this cooling trend, Yunnan has also been affected by an aridification since the Miocene. Palaeoclimate reconstructions indicate that environments were humid in Yunnan during the late Miocene, and thereafter a drying trend occurred(Xia et al., 2009; Jacques et al., 2011; Sun et al., 2011; Xing et al., 2012). For example, the late Miocene Xianfeng flora revealed a mean annual precipitation(MAP)of 1206e1613 mm, higher than the presentday value of 1011 mm(Xing et al., 2012). This post-Miocene aridification appears to be particularly pronounced in the dry season. Various palaeoclimate reconstructions indicate that Yunnan had higher precipitation in summer but less rainfall in winter during the late Miocene(Xia et al., 2009; Jacques et al., 2011; Xing et al., 2012)and late Pliocene(Xie et al., 2012; Su et al., 2013a; Huang et al., 2015a). For instance, during the late Miocene the mean monthly precipitation of the three wettest months was 150e250 mm, while that of the three driest months was less than 50 mm, suggesting a prominent precipitation seasonality. These palaeoclimate results also demonstrate that summer precipitation has increased while winter precipitation has decreased since the late Miocene(Jacques et al., 2011; Xing et al., 2012; Su et al., 2013a; Huang et al., 2015a), suggesting a strengthening of the precipitation seasonality in Yunnan.
8. ConclusionBased on our integration of published palaeobotanical data from Yunnan, our summary reaches six key points as stated below.
(1) Yunnan has a rich Cenozoic plant fossil record which has been investigated extensively, particularly over the past two decades, with an increasing number of fossil taxa being reported. These fossil taxa are important for answering past plant diversity and floral evolution in this region.
(2) Plant species from the Cenozoic of Yunnan are diverse. To date, approximately 386 fossil species belonging to 170 genera of 66 families have been documented. They cover ferns, gymnosperms and angiosperms, of which angiosperms are by far the most diverse, including 353 species grouped into 155 genera of 60 families. In the fossil angiosperms, Fagaceae, Fabaceae, Juglandaceae, Lauraceae and Rosaceae are among the most diverse families; and Querucs, Castanopsis, Acer, Lithocarpus, Cyclobalanopsis and Cinnamomum are among the most species-rich genera.
(3) Most of the families and genera represented in the fossil record are still part of the modern natural vegetation in Yunnan, but some genera have disappeared, possibly as a result of an aridification and/or extension of the dry season. So far, seven genera are known to have become extinct in Yunnan, namely Berryophyllum, Cedrelospermum, Cedrus, Palaeocarya, Podocarpium, Sequoia and Wataria.
(4) In the late Miocene, Yunnan had three floristic regions: a northern subtropical zone in the northeast, a subtropical zone in the east, and a tropical zone in the southwest. In the late Pliocene, Yunnan had two kinds of floristic regions: a subalpine floristic region in the northwest, and two subtropical floristic regions separately in the southwest and in the eastern center. Floristic characters changed significantly from the late Miocene to late Pliocene through to today, presumably attributed to altitude changes.
(5) In the late Miocene, southern Yunnan had more elements of the Pantropic type than northern Yunnan, with the southwest being obviously the richest in the Pantropic type, in agreement with results from floristic analyses. Three areal types including the North Temperate type, the Tropical Asia and Tropical America disjunct type, and the East Asia and North America disjunct type, were common in most of Yunnan except for the northeast. In the Pliocene, vegetation of Yunnan was dominated by the North Temperate type, but southwestern Yunnan also had abundant genera of the Tropical Asia (Indo-Malaysia) type and the East Asia and North America disjunct type.
(6) In the Neogene, Yunnan was warmer and more humid than today. Since the late Miocene, temperature has declined significantly in northern Yunnan but has remained almost at the same level in southern Yunnan. Summer precipitation has increased while winter rainfall has decreased, suggesting an intensification of the precipitation seasonality to the present.
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.pld.2016.11.004.
AcknowledgementsThis study was supported by the National Natural Science Foundation of China (No. U1502231), the Foundation of the State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (No. 163108), and the foundation of the Yunnan Key Laboratory for Palaeobiology, Yunnan University (No. 2015DG007-KF01). We thank Dr. Shu-Feng Li (Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences) for help with this study; and professor Robert A. Spicer (The Open University) and the other anonymous reviewer for constructive comments in improving the manuscript. This work is a contribution to Neogene Climate Evolution in Eurasia (NECLIME).
Chang L, Guo Z, Deng C, Wu H, Ji X, Zhao Y, Zhang C, Ge J, Wu B, Sun L, Zhu R, 2015. Pollen evidence of the palaeoenvironments of Lufengpithecus lufengensis in the Zhaotong basin,southeastern margin of the Tibetan plateau. Palaeogeogr. Palaeoclimatol. Palaeoecol., 435: 95-104. DOI:10.1016/j.palaeo.2015.06.007 |
Clark M.K, House M.A, Royden L.H, Whipple K.X, Burchfiel B.C, Zhang X, Tang W, 2005. Late cenozoic uplift of southeastern tibet. Geology, 33: 525-528. DOI:10.1130/G21265.1 |
Colani M.M, 1920. E'tude sur les flores Tertiares de quelques gisments de lignit de 1'Indochine et du Yunnan. Bull. Serv. Geol. 1'Indochine, 8: 1-260. |
Forman L.L, 1966. Generic delimitation in the castaneoideae. (Fagaceae). Kew Bull., 18: 421-426. DOI:10.2307/4115795 |
Ge H.R, Li D.Y, 1999. Cenozoic coal-bearing basins and coal-forming regularity in West Yunnan. Kunming: Yunnan Science and Technology Press.
|
Guo S.X, 2011. The late Miocene Bangmai flora from Lincang county of Yunnan,southwestern China. Acta Palaeontol. Sin., 50: 353-408. |
Guo S.X, Zhang G.F, 2002. Oligocene sanhe flora in Longjing county of Jilin,northeast China. Acta Palaeontol. Sin., 41: 193-210. |
Guo, S.X., Zhou, Z.K., 1992. The mega fossil legumes from China. In: Herendeen, P.S., Dilcher, D.L. (Eds.), Advances in Legume Systematics: Part 4, the Fossil Record. Royal Botanic Gardens, Kew, London, pp. 207-223.
|
Hably L, Thiebaut M, 2002. Revision of Cedrelospermum (Ulmaceae) fruits and leaves from the tertiary of Hungary and France. Palaeontogr. Abt. B 262, 262: 71-90. |
Hjelmquist H, 1948. Studies on the floral morphology and phylogeny of the Amentiferae. Bot. Not, 2: S1-S171. |
Hu, Q., 2013. Leaf Morphological Evolution of Quercus Delavayi Complex and Cupule Morphology of Quercus Subg. Cyclobalanopsis. Master's Thesis. Kunming Institute of Botany, Chinese Academy of Sciences.
|
Hu Q, Xing Y, Hu J, Huang Y, Ma H, Zhou Z, 2014. Evolution of stomatal and trichome density of the Quercus delavayi complex since the late Miocene. Chin. Sci. Bull., 59: 310-319. DOI:10.1007/s11434-013-0038-z |
Huang, C., Zhang, Y., Bartholomew, B., 1999. Fagaceae. In: Flora of China Editorial Committee (Eds.), Flora of China. Vol. 4 (Cycadaceae through Fagaceae). Science Press, Beijing, pp. 314-402. Missouri Botanical Garden Press, St. Louis.
|
Huang J, Su T, Lebereton-Anberree J, Zhang S.-T, Zhou Z.-K, 2016a. The oldest Mahonia (Berberidaceae) fossil from East Asia and its biogeographic implications. J. Plant Res., 129: 209-223. DOI:10.1007/s10265-015-0775-y |
Huang, Y.-J., 2012. The Late Pliocene Fudong Flora from Lanping, Yunnan, and the Neoegene Climates in Hengduan Mountains. Ph.D. Thesis. Kunming Institute of Botany, Chinese Academy of Sciences.
|
Huang Y.-J, Chen W.-Y, Jacques F.M.B, Liu Y.-S, Utescher T, Su T, Ferguson D.K, Zhou Z.-K, 2015a. Late Pliocene temperatures and their spatial variation at the southeastern border of the Qinghai-Tibet Plateau. J. Asian Earth Sci., 111: 44-53. DOI:10.1016/j.jseaes.2015.04.048 |
Huang Y.-J, Jacques F.M.B, Liu Y.-S, Su T, Xing Y, Xiao X, Zhou Z, 2012. New fossil endocarps of Sambucus (Adoxaceae) from the upper Pliocene in SW China. Rev. Palaeobot. Palynol., 171: 152-163. DOI:10.1016/j.revpalbo.2011.11.008 |
Huang Y.-J, Ji X, Su T, Wang L, Deng C, Li W, Luo H, Zhou Z, 2015b. Fossil seeds of Euryale (Nymphaeaceae) indicate a lake or swamp environment in the late Miocene Zhaotong Basin of southwestern China. Sci. Bull., 60: 1768-1777. DOI:10.1007/s11434-015-0870-4 |
Huang Y.-J, Liu Y.-S, Jacques F.M.B, Su T, Xing Y.-W, Zhou Z.-K, 2013. First discovery of Cucubalus (Caryophyllaceae) fossil,and its biogeographical and ecological implications. Rev. Palaeobot. Palynol., 190: 41-47. DOI:10.1016/j.revpalbo.2012.11.011 |
Huang Y.-J, Su T, Zhou Z.-K, 2016b. Late Pliocene diversity and distribution of Drynaria (Polypodiaceae) in western Yunnan explained byforest vegetation and humid climates. Plant Divers, 38: 194-200. DOI:10.1016/j.pld.2016.06.003 |
Jacques F.M.B, Guo S.-X, Su T, Xing Y.-W, Huang Y.-J, Liu Y.-S, Ferguson D.K, Zhou Z.-K, 2011. Quantitative reconstruction of the Late Miocene monsoon climates of southwest China: a case study of the Lincang flora from Yunnan Province. Palaeogeogr. Palaeoclimatol. Palaeoecol., 304: 318-327. DOI:10.1016/j.palaeo.2010.04.014 |
Jacques F.M.B, Su T, Spicer R.A, Xing Y.-W, Huang Y.-J, Zhou Z.-K, 2014. Late Miocene southwestern Chinese floristic diversity shaped by the southeastern uplift of the Tibetan Plateau. Palaeogeogr. Palaeoclimatol. Palaeoecol., 411: 208-215. DOI:10.1016/j.palaeo.2014.05.041 |
Jacques F.M.B, Su T, Zhou Z.-K, 2013. The first fossil Microsoroid fern (Palae- osorum ellipticum gen. et sp. nov.) from the middle Miocene of Yunnan,SW China. J. Syst. Evol., 51: 758-764. |
Jähnichen H, Friederick W.L, Takac M, 1984. Engelhardioid leaves and fruits from the European Tertiary,part II. Tert. Res., 6: 109-134. |
Jia L.-B, Manchester S.R, Su T, Xing Y.-W, Chen W.-Y, Huang Y.-J, Zhou Z.-K, 2015. First occurrence of Cedrelospermum (Ulmaceae) in Asia and its biogeographic implications. J. Plant Res., 128: 747-761. DOI:10.1007/s10265-015-0739-2 |
Jin J, 2009. Two Eocene fossil fruits from the changchang basin of Hainan Island,China. Rev. Palaeobot. Palynol., 153: 150-152. DOI:10.1016/j.revpalbo.2008.07.010 |
Jones J.H, Dilcher D.L, 1988. A study of the "Dryphyllum" leaf forms from the Paleogene of southeastern North America. Palaeontogr. Abt. B, 208: 53-80. |
Kumar R, Singh C, Malik S, Ali A, Nayital R.K, 2011. Effect of storage conditions on germinability of Himalayan Cedar (Cedrus deodara Roxb, G. Don) seeds. Indian For, 137: 1099-1102. |
Lebreton-Anberrée J, Li S, Shu-Feng L, Spicer R.A, Zhang S.-T, Su T, Deng C, Zhou Z.-K, 2016. Lake geochemistry reveals marked environmental change in southwest China during the mid miocene climatic optimum. Sci. Bull., 61: 897-910. DOI:10.1007/s11434-016-1095-x |
Li, Y.-J., 2015. System Classification of the Fossil Woods of the Miocene from Shengli Coal Mine, Pu'er, Yunnan Province. Master's Thesis. Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences.
|
Li Y.-J, Oskolski A.A, Jacques F.M.B, Zhou Z.-K, 2015. New middle Miocene fossil wood of Wataria (Malvaceae) from southwest China. IAWA J., 36: 345-357. DOI:10.1163/22941932-20150105 |
Liang X.-Q, Ferguson D.K, Jacques F.M.B, Su T, Wang L, Zhou Z.-K, 2016. A new Celastrus species from the middle miocene of Yunnan,China and its palae- oclimatic and palaeobiogeographic implications. Rev. Palaeobot. Palynol., 225: 43-52. DOI:10.1016/j.revpalbo.2015.11.005 |
López-Pujol J, Zhang F.-M, Ge S, 2006. Plant biodiversity in China: richly varied,endangered and in need of conservation. Biodivers. Conserv., 15: 3983-4026. DOI:10.1007/s10531-005-3015-2 |
López-Pujol J, Zhang F.-M, Sun H.-Q, Ying T.-S, Ge S, 2011. Centres of plant endemism in China: places for survival or for speciation?. J. Biogeogr., 38: 1267-1280. DOI:10.1111/jbi.2011.38.issue-7 |
Maheshwari, P., Biswas, C., 1970. Cedrus (Botanical Monograoh No. 5). Council of Scientific and Industrial Research, Rafi Marg, New Delhi.
|
Manchester S.R, 1987. Extinct ulmaceous fruits from the tertiary of Europe and western North America. Rev. Palaeobot. Palynol., 52: 119-129. DOI:10.1016/0034-6667(87)90049-2 |
Manchester S.R, 1989. Attached reproductive and vegetative remains of the extinct American-European genus Cedrelospermum (Ulmaceae) from the early Tertiary of Utah and Colorado. Am. J. Bot., 76: 256-276. DOI:10.2307/2444668 |
Manchester S.R, 1999. Biogeographical relationship of North America tertiary floras. Ann. Mo. Bot. Gard., 86: 472-522. DOI:10.2307/2666183 |
Manchester S.R, Tiffney B.H, 2001. Integration of paleobotanical and neobotanical data in the assessment of phytogeographic history of holarctic angiosperm clades. Int.J. Plant Sci, 162: S19-S27. DOI:10.1086/323657 |
Manos P, Zhou Z.K, Cannon C.H, 2001. Systematics of Fagaceae: phylogenetic tests of reproductive trait evolution. Int. J. Plant Sci., 162: 1361-1379. DOI:10.1086/322949 |
Meng H.-H, Jacques, Su T, Huang Y.-J, Zhang S.-T, Ma H.-J, Zhou Z.-K, 2014. New biogeographic insight into Bauhinia s.l.. (Leguminosae): integration from fossil records and molecular analyses. BMC Evol. Biol. 14, 14: 181. |
Meng H.-H, Su T, Huang Y.-J, Zhu H, Zhou Z.-K, 2015. Late Miocene Palaeocarya (Engelhardieae: Juglandaceae) from southwest China and its biogeographic implications. J. Syst. Evol., 53: 499-511. DOI:10.1111/jse.v53.6 |
Olson, D.F., Roy, D.F., Walters, G.A., 1990. Sequoia sempervirens (D. Don) Endl. Redwood. In: Burns, R.M., Honkala, B.H. (Eds.), Silvics of North America, Vol. 1, Conifers. Agriculture Handbook, vol. 654. USDA Forest Service, Washington D.C, pp. 541-551.
|
Ruth S, Renee M, Li H, Fang Z, Wang Y, 2008. Spatial patterns of plant diversity and communities in Alpine ecosystems of the Hengduan Mountains,northwest Yunnan,China. J. Plant Ecol., 1: 117-136. DOI:10.1093/jpe/rtn012 |
Su, T., 2010. On the Establishment of the Leaf Physiognomy-climate Model and a Study of the Late Pliocene Yangjie Flora, Southwest China. Ph.D. Thesis. Kunming Institute of Botany, Chinese Academy of Sciences.
|
Su T, Huang Y.-J, Meng J, Zhang S.-T, Huang J, Zhou Z.-K, 2016. A Miocene leaf fossil record of Rosa (R. fortuita n. sp.) from its modern diversity center in SW China.. Palaeoworld, 25: 104-115. DOI:10.1016/j.palwor.2015.05.010 |
Su T, Jacques F.M.B, Liu Y.-S, Xiang J.-Y., Xing Y.-W, Huang Y.-J, Zhou Z.-K, 2011. A new Drynaria (Polypodiaceae) from the upper Pliocene of southwest China. Rev. Palaeobot. Palynol., 164: 132-142. DOI:10.1016/j.revpalbo.2010.11.011 |
Su T, Jacques F.M.B, Spicer R.A, Liu Y.-S, Huang Y.-J, Xing Y.-W, Zhou Z.-K, 2013a. Post-Pliocene establishment of the present monsoonal climate in SW China: evidence from the late Pliocene Longmen megaflora. Clim. Past., 9: 1911-1920. DOI:10.5194/cp-9-1911-2013 |
Su T, Liu Y.-S, Jacques F.M.B, Huang Y.-J, Xing Y.-W, Zhou Z.-K, 2013b. The intensification of the East Asian winter monsoon contributed to the disappearance of Cedrus (Pinaceae) in southwestern China. Quat. Res., 80: 316-325. DOI:10.1016/j.yqres.2013.07.001 |
Sun B.-N, Wu J.-Y, L iu, (Christopher) Y.-S, Ding S.-T, Li X.-C, Xie S.-P, Yan D.-F, Lin Z.-C, 2011. Reconstructing Neogene vegetation and climates to infer tectonic uplift in western Yunnan,China. Palaeogeogr. Palaeoclimatol. Palaeoecol., 304: 328-336. DOI:10.1016/j.palaeo.2010.09.023 |
Tanai T, Uemura K, 1983. Engelhardia fruits from the tertiary of Japan. Hokkaido University J. Fac. Sci., 22: 249-260. |
Tao, J.R.,1986. Neogene flora of Fudong and its significance in middle watershed of Selween-Mekong-Yangtze Rivers. In: Tao, J.R. (Ed.), Hengduan Mountain Investigation Special. Science and Technology Publishing House, Beijing, pp. 58-65.
|
Tao, J.R., Chen, M.H., 1983. Neogene flora of south part of the watershed of Salweed- Mekong-Yangtze river (the Linczan region) Yunnan. In: Exploration of Heng- duan Mountain Area, vol. 1. Beijing Publishing House of Science and Technology, Beijing, pp. 74-89.
|
Terada K, Suzuki M, 1998. Revision of the so-called Reevesia fossil woods from the Tertiary in Japan - a proposal of the new genus Wataria (Sterculiaceae). Rev. Palaeobot. Palynol., 103: 235-251. DOI:10.1016/S0034-6667(98)00039-6 |
Tong G.-B, Bai S.-Y, Zhou S.-J, Zhang Z.-P, 1990. The characteristics of the pollen spore time series fluctuation and its significance in the geological environments of the Quaternary in Kunming Basin. Acta Bot. Sin., 32: 146-156. |
Turkington R, Harrower W.L, 2016. An experimental approach to addressing ecological questions related to the conservation of plant biodiversity in China. Plant Divers, 38: 1-10. DOI:10.1016/j.pld.2016.05.005 |
Uhl D, Klotz S, Traiser C, Thiel C, Utescher T, Kowalski E, Dilcher D.L, 2007. Cenozoic paleotemperatures and leaf physiognomy—a European perspective. Palaeogeogr. Palaeoclimatol. Palaeoecol., 248: 24-31. DOI:10.1016/j.palaeo.2006.11.005 |
Utescher T, Bruch A.A, Micheels A, Mosbrugger V, Popova S, 2011. Cenozoic climate gradients in Eurasia—a palaeo-perspective on future climate change?. Palaeogeogr. Palaeoclimatol. Palaeoecol, 304: 351-358. DOI:10.1016/j.palaeo.2010.09.031 |
Wang A, Yang M, Liu J, 2005. Molecular phylogeny,recent radiation and evolution of gross morphology of the rhubarb genus Rheum (Polygonaceae) inferred from chloroplast DNA trn L-F sequences. Ann. Bot., 96: 489-498. DOI:10.1093/aob/mci201 |
Wang, H.-B., 2016. Fossil woods from Miocene Xundian County, Yunnan and Their Palaeoenvironmental Implications. Ph.D Thesis. Kunming Institute of Botany, Chinese Academy of Sciences.
|
Wang, H.-B., Oskolski, A.A., Jacques, F.M.B., Wang, Y.-H., Zhou, Z.-K., in press. Lignified woods of Pinus (Pinaceae) from the late Miocene of central Yunnan, China, and their biogeographic and paleoclimatic implications. Palaeoworld.
|
Wang L, Jacques F.M.B, Su T, Xing Y, Zhang S, Zhou Z, 2013. The earliest fossil bamboos of China (middle Miocene,Yunnan) and their biogeographical importance. Rev. Palaeobot. Palynol., 197: 253-265. DOI:10.1016/j.revpalbo.2013.06.004 |
Wang W.-M, Shu J.-W, 2004. Late cenozoic palynofloras from Qujing basin,Yunnan,China. Acta Palaeontol. Sin., 43: 254-261. |
Waston, F.D., Eckenwalder, J.E., 1993. Cupressaceae. In: Flora of North America Editorial Committee (Ed.), Flora of North America, North of Mexico, vol. 2. Oxford University Press, Oxford, pp. 399-422.
|
Wen W.-W, Xie S.-P, Liu K.-N, Sun B.-N, Wang L, Li H, Dao K.-Q, 2013. Two species of fern macrofossil from the late Miocene ofLincang,Yunnan,China and their paleoecological implications. Palaeoworld, 22: 144-152. DOI:10.1016/j.palwor.2013.06.004 |
Writing Group of Cenozoic Plants in China (WGCPC), 1978. Fossil Plants in China,Vol. 3. Beijing: Cenozoic Plants in China. Science Press.
|
Writing Group of Yunnan Vegetation (WGYV), 1987. Vegetation of Yunnan. Beijing: Science Press.
|
Wu, J.-Y., 2010. The Pliocene Tuantian Flora of Tengchong, Yunnan Province and its Paleoenvironmental Analysis. Ph.D. Thesis. Lanzhou University.
|
Wu J.-Y, Sun B.-N, Xie S.-P, Ding S.-T, Wen W.-W, 2012. Dimorphic fronds and in situ spores of Drynaria (Polypodiaceae) from the upper Pliocene of southwest China. Rev. Palaeobot. Palynol., 172: 1-9. DOI:10.1016/j.revpalbo.2012.01.007 |
Wu Z.Y, 1965. The tropical floristic affinity of the flora of China. Chin. Sci. Bull., 1: 25-33. |
Wu Z.Y, 1979. Flora of Yunnan,vol. 2. Beijing: Science Press.
|
Wu Z.Y, 1988. Hengduan Mountain flora and her significance. J. Jap. Bot., 63: 297-311. |
Wu Z.Y, 1991. The areal-types of Chinese genera of seed plants. Acta Bot. Yunnanica, 4: S1-S139. |
Wu Z.Y, Zhou Z.-K, Li D.-Z, Peng H, Sun H, 2003. The areal-types of the world families of seed plants. Acta Bot. Yunnanica, 25: 245-257. |
Xia K, Su T, Liu Y.-S, Xing Y.-W, Jacques F.M.B, Zhou Z.-K, 2009. Quantitative climate reconstructions of the late Miocene Xiaolongtan megaflora from Yunnan,southwest China. Palaeogeogr. Palaeoclimatol. Palaeoecol., 276: 80-86. DOI:10.1016/j.palaeo.2009.02.024 |
Xiao X.Y, Shen J, Wang S.M, Xiao H.F, Tong G.B, 2010. The variation of the southwest monsoon from the high resolution pollen record in Nanbanbang Basin,Yunnan Province,China for the last 2.78 Ma. Palaeogeogr. Palaeoclimatol. Palaeoecol., 287: 45-57. DOI:10.1016/j.palaeo.2010.01.013 |
Xie, S., 2007. Numerical Taxonomy of Winged Fruits and Paleoenvironmental Reconstruction Based on Angiosperm Leaves from the Neogene of West Yunnan. Ph. D. Thesis. Lanzhou University.
|
Xie S, Manchester S.R, Liu K, Wang Y, Shao Y, 2014. Firmiana (Malvaceae: Sterculioideae) fruits from the upper Miocene of Yunnan,southwest China. Geobios, 47: 271-279. DOI:10.1016/j.geobios.2014.03.005 |
Xie S, Sun B, Dilcher D.L, Yan D, Wu J, Lin Z, 2010. Numerical taxonomy of Palaeocarya (Juglandaceae) from the Mangbang formation of west Yunnan,China. Rev. Palaeobot. Palynol., 162: 193-202. DOI:10.1016/j.revpalbo.2010.07.001 |
Xie S, Sun B, Wu J, Lin Z, Yan D, Xiao L, 2012. Palaeoclimatic estimates for the late Pliocene based on leaf physiognomy from western Yunnan,China. Turk. J. Earth Sci., 21: 251-261. |
Xing, Y., 2010. The Late Miocene Xianfeng Flora, Yunnan, Southwest China and its Quantitative Palaeoclimatic Reconstructions. Ph.D. Thesis. Kunming Institute of Botany, Chinese Academy of Sciences.
|
Xing Y, Liu Y.-S, Su T, Jacques F.M.B, Zhou Z, 2010. Pinus prekesiya sp. nov. from the upper Miocene of Yunnan,southwestern China and its biogeographical implications. Rev. Palaeobot. Palynol., 160: 1-9. |
Xing Y, Liu Y.-S, Su T, Jacques F.M.B, Zhou Z.-K, 2013. A new Tsuga species from the upper Miocene of Yunnan,southwestern China and its palaeogeographic significance. Palaeoworld, 22: 159-167. DOI:10.1016/j.palwor.2013.09.003 |
Xing Y, Utescher T, Jacques F.M.B, Su T, Liu Y.-S, Huang Y, Zhou Z, 2012. Pa- leoclimatic estimation reveals a weak winter monsoon in southwestern China during the late Miocene: evidence from plant macrofossils. Palaeogeogr. Palaeoclimatol. Palaeoecol. 358-, 360: 19-26. |
Xu Q, Qiu J, Zhou Z, Jin J, 2015. Eocene Podocarpium (Leguminosae) from south China and its biogeographic implications. Front. Plant Sci, 6: 938. |
Yu W.-B, Liu M.-L, Wang H, Mill R.R, Ree R.H, Yang J.-B., Li D.-Z, 2015. Towards a comprehensive phylogeny of the large temperate genus Pedicularis (Oroban- chaceae),with an emphasis on species from the Himalaya-Hengduan Mountains. BMC Plant Biol, 15: 176. DOI:10.1186/s12870-015-0547-9 |
Zhang J.-J, Ding L, 2003. East-west extension in Tibetan Plateau and its significance to tectonic evolution. Chin. J. Geol., 38: 179-189. |
Zhang J.-W, D'Rozario A, Adams J.M, Li Y, Liang X.-Q, Jacques F.M.B, Su T, Zhou Z.-K, 2015a. Sequoia maguanensis,a new Miocene relative of the coast redwood,Sequoia sempervirens,from China: implications for paleogeography and paleoclimate. Am. J. Bot., 102: 103-118. DOI:10.3732/ajb.1400347 |
Zhang J.-W, Huang J, D'Rozario A, Adams J.M, Zhou Z.-K, 2015b. Calocedrus shengxianensis,a late Miocene relative of C. macrolepis (Cupressaceae) from South China: implications for paleoclimate and evolution of the genus. Rev. Palaeobot. Palynol., 222: 1-15. |
Zhao, L.-C., 2002. Fruits and Seeds from Yunnan in the Age of Miocene and from Shanxi in the Age of Pliocene. Ph.D. Thesis. Institute of Botany, Chinese Academy of Sciences.
|
Zhao L.-C, Wang Y.-F, Liu C.-J, Li C.-S, 2004. Climatic implications of fruit and seed assemblages from Miocene of Yunnan,southwestern China. Quatern. Int, 117: 81-89. DOI:10.1016/S1040-6182(03)00118-6 |
Zhou Z, 1996. Studies on Dryophyllum complex from China and its geological and systematic implications. Acta Bot. Sin., 38: 666-671. |
Zhu H, 2013. Geographical elements of seed plants suggest the boundary of the tropical zone in China. Palaeogeogr. Palaeoclimatol. Palaeoecol., 386: 16-22. DOI:10.1016/j.palaeo.2013.04.007 |
Zhu, H., 2016. The Late Pliocene Nanbanbang Carpoflora from Heqing, Yunnan, and its Paleoenvironment. Ph.D. Thesis. Kunming Institute of Botany, Chinese Academy of Sciences.
|
Zhu H, Huang Y.-J, Su T, Zhou Z.-K, 2016. New fossil seeds of Eurya (Theaceae) from East Asia and their paleobiogeographic implications. Plant Divers, 38: 125-132. DOI:10.1016/j.pld.2016.05.001 |
Zhu H, Jacques F.M.B, Wang L, Xiao X.-H, Huang Y.-J, Zhou Z.-K, 2015. Fossil endocarps of Aralia (Araliaceae) from the upper Pliocene of Yunnan in southwest China,and their biogeographical implications. Rev. Palaeobot. Palynol., 223: 94-103. DOI:10.1016/j.revpalbo.2015.09.010 |