 2016, Vol. 42
2016, Vol. 42扩展功能
文章信息
- 王朝义, 王强, 郑曰勇, 李聪, 郭敦伟, 唐华
- WANG Chaoyi, WANG Qiang, ZHENG Yueyong, LI Cong, GUO Dunwei, TANG Hua
- 吉林大学学报(医学版), 2016, 42(04): 653-658
- Journal of Jilin University (Medicine Edition), 2016, 42(04): 653-658
- 10.13481/j.1671-587x.20160404
-
文章历史
- 收稿日期: 2015-06-07




2. 重庆医科大学附属第一医院妇科, 重庆 400016
2. Department of Gynaecology, First Affiliated Hospital, Chongqing Medical University, Chongqing 400016, China
肿瘤恶病质(cancer cachexia,CC)是一种多因子综合征,主要表现为进行性骨骼肌组织减少,并导致进一步功能损害[1]。肿瘤患者恶病质的发生率非常高,约50%晚期肿瘤和80%终末期肿瘤患者发生恶病质。恶病质对肿瘤患者放、化疗的敏感性及耐受性、生活质量和生存期均有严重影响[2]。但目前肿瘤恶病质的具体发生机制尚不完全清楚,亦缺乏有效的防治措施。骨骼肌消耗是促进肿瘤恶病质发生发展的关键环节,近年来研究[3, 4, 5]显示:肌肉生长抑制素(myostatin,Mstn)在肿瘤恶病质的发生发展中发挥重要作用。肺癌转移相关转录本1 (metastasis-associated lung adenocarcinoma transcript 1,MALAT1)是一种高度保守并具有多种功能的长链非编码RNA(long non-coding RNA,LncRNA)。Watts等[6]发现:在Mstn介导的骨骼肌消耗动物模型中LncRNA-MALAT1表达水平明显下降,LncRNA-MALAT1可能与骨骼肌消耗有关联。进一步研究[7]显示:LncRNA-MALAT1可抑制其靶基因Caspase-3的表达。而Caspase-3在肿瘤恶病质骨骼肌降解过程中发挥着启动作用,并能与其他的蛋白水解酶协同作用加速蛋白降解[8]。在肿瘤恶病质中,LncRNA-MALAT1的作用及其与Mstn和Caspase-3的关系鲜有报道。 卵泡抑素(follistatin, FST)是一种有效的Mstn抑制剂,能够阻断Mstn对骨骼肌的负性调控作用[9]。因此本研究旨在探讨FST能否通过抑制Mstn表达,上调LncRNA-MALAT1表达,进而抑制Caspase-3表达,达到缓解肿瘤恶病质小鼠骨骼肌消耗的目的。
1 材料与方法 1.1 实验动物及主要试剂32只清洁级BALB/c小鼠,健康雄性,7~9周龄,体质量22~26 g,购自重庆医科大学实验动物中心,动物合格证号:SYXK(渝)2012-0001。小鼠结肠腺癌C26细胞株由重庆医科大学附属第一医院实验研究中心提供。FST购于美国R & D公司,Caspase-3一抗购于英国Abcam公司,Mstn和β-actin一抗及相应二抗购于美国Proteintech公司,Western blotting相关试剂盒购自广州碧云天生物有限公司,Real-time PCR试剂盒购自日本TaKaRa公司。
1.2 实验分组及小鼠恶病质模型的建立32只BALB/c小鼠随机分为健康对照组(HC组)、FST预防组(FP组)、FST治疗组(FT组)和肿瘤恶病质组(CC组),每组8只。小鼠适应性喂养1周后,取对数生长期的C26细胞接种于FP、FT和CC组小鼠左侧腋窝皮下(约1×106个细胞/只),HC组小鼠接种等体积PBS。荷瘤小鼠出现精神萎靡,毛发粗糙、泛黄并失去光泽,自发性活动减少等表现,且体质量下降与健康对照组比较差异有统计学意义时,即表明小鼠进入恶病质状态[10]。
1.3 指标监测与分组处理每日定时观察小鼠毛发、精神状况、体质量及肿瘤生长情况。肿瘤体积按公式 V=ab2/2计算,a表示长度,b表示宽度。于接种后第6天(可触及肿瘤结节后)开始给予FP组小鼠FST(5 μg·kg-1·d-1)腹腔注射,于接种后第12天(小鼠进入恶病质状态后)开始给予FT组小鼠FST(5 μg·kg-1·d-1)腹腔注射,HC组和CC组小鼠以等量生理盐水代替。采用ZH-ZFT型视频记录分析系统检测小鼠1h内自发性活动总路程。第20天采用戊巴比妥钠麻醉小鼠,摘除眼球后取血,随即取少量血液采用血糖检测仪测定血糖水平,其余血液4℃静置2h,4℃、4200r·min-1离心15min,取上清液,自动生化仪检测血清总蛋白和白蛋白水平。剥离肿瘤和腓肠肌组织并称质量,左侧腓肠肌组织置于4%多聚甲醛中固定,肿瘤及右侧腓肠肌组织保存于液氮中备用。
1.4 小鼠腓肠肌纤维横切面积检测腓肠肌组织固定48 h,常规石蜡包埋。将组织块按肌纤维横切面方向行连续切片。石蜡切片常规方法脱蜡,依次行苏木素染色、1%盐水酒精分化、伊红染色和梯度乙醇脱水,中性树脂封片。每个样本取5张切片,每张切片取3个视野,于400倍光学显微镜下采集图片,采用Image Pro Plus6.0软件计算腓肠肌纤维横切面面积,取平均值。
1.5 Real-time PCR法检测小鼠不同组织中Mstn、LncRNA-MALAT1和Caspase-3mRNA表达水平取50mg腓肠肌组织或肿瘤组织,加入1mL Trizol提取组织总RNA,测纯度和浓度后取1 μg RNA反转录合成cDNA。按SYBROR Premix Ex TaqTMⅡ试剂盒说明书以β-actin为内参建立10 μL反应体系。反应 条件:95℃预变性3 min,95℃变性5s,58℃退火15s,72 ℃延伸30s,循环40次。Mstn引物:F为5′-ACTGGAATCCGATCTCTGAAACTT-3′,R为5′-TCCTGGT- CCTGGGAAGGTTAC-3′,177bp;LncRNA-MALAT1引物:F为5′-CTCACTAAAGGCACTGAAGGCT-3′,R为5′-CACTGCTAATCAAAA-CCGACCA -3′,120bp;Caspase-3引物:F为5′-TGGGACTGATGAGGAGATGGC -3′,R为5′-TGCTGCAAAGGGACTGGATG -3′,137bp;β-actin引物:F为5′- AGATTACTGCTCTGGCTCCTAGC-3′,R为5′-ACTCATCGTACTCCTGCTTGCT-3′,147 bp。PCR扩增反应结束后以β-actin为内参,采用公式2-△△ct计算各基因mRNA表达水平。
1.6 Western blotting法检测小鼠不同组织中Mstn和Caspase-3蛋白表达水平取50mg腓肠肌或肿瘤组织,加入350 μL RIPA/PMSF混合液裂解,提取总蛋白,采用BCA蛋白浓度测定试剂盒测定浓度。10%SDS-PAGE凝胶电泳分离,电转至PVDF膜上,5%脱脂奶粉于37℃条件下封闭90 min,然后与Mstn(1∶1000)、Caspase-3(1∶1 500)和β-actin(1∶1 000)一抗4℃孵育过夜,洗膜,再与相应二抗孵育,洗膜后化学发光法显影。结果采用凝胶成像/图像分析系统处理,以目的蛋白条带灰度值与β-actin条带灰度值的比值作为目的蛋白相对表达水平。
1.7 统计学分析
采用SPSS17.0统计软件进行统计学分析。小鼠体质量,肿瘤质量和体积,自发性活动总路程,血清生化指标,腓肠肌质量,腓肠肌肌纤维横切面面积,Mstn、LncRNA-MALAT1和Caspase-3蛋白及mRNA表达水平均以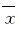 ±s表示,多组间比较采用单因数方差分析,两组间均值比较采用SNK-q检验。以P<0.05为差异有统计学意义。
±s表示,多组间比较采用单因数方差分析,两组间均值比较采用SNK-q检验。以P<0.05为差异有统计学意义。
C26细胞接种前各组小鼠体质量比较差异无统计学意义(P>0.05)。接种后第6天,荷瘤小鼠皮下可触及肿瘤结节。第12天,与HC组比较,FT组和CC组小鼠体质量及自发性活动总路程明显降低(P<0.05),并出现毛发粗糙、失去光泽和精神萎靡等恶病质表现;FP组小鼠进入恶病质状态的时间明显延后。第20天,与CC组比较,FP组和FT组小鼠经FST干预后体质量下降幅度减少(P<0.05),自发性活动总路程增加(P<0.05),且FP组大于FT组(P<0.05)。与CC组比较,FP组小鼠肿瘤质量和体积均下降(P<0.05),而FT组差异无统计学意义 (P>0.05)。见表1和表2。
| (n=8,±s) | |||||
| Group | Body weight(m/g) | Tumor weight(m/g) | Tumor volume(V/cm3) | ||
| (t/d) 1 | 12 | 20 | |||
| HC | 23.63±1.57 | 26.40±1.18 | 30.06±1.27 | - | - |
| CC | 24.36±2.05 | 24.11±0.75 * | 20.23±1.01 * | 4.03±0.83 | 3.40±0.45 |
| FP | 24.26±1.96 | 25.80±0.66 | 24.67±1.29 * △# | 2.37±0.70 △ | 2.03±0.07 △ |
| FT | 24.83±2.59 | 24.10±0.65 * | 22.23±0.61 * △ | 3.16±0.32 | 2.65±0.61 |
| “-”:No data. * P<0.05 compared with HC group; △ P<0.05 compared with CC group; # P<0.05 compared with FT group. | |||||
| (n=8,±s,l/m) | |||
| Group | Distance of spontaneous activity | ||
| (t/d) 1 | 12 | 20 | |
| HC | 408.56±64.69 | 426.77±55.99 | 421.47±26.78 |
| CC | 409.84±52.26 | 306.73±46.51 * | 152.30±34.82 * |
| FP | 417.33±57.03 | 392.00±48.96 | 329.66±66.75 * △# |
| FT | 418.91±44.73 | 312.91±44.25 * | 213.53±30.51 * △ |
| * P<0.05 compared with HC group; △ P<0.05 compared with CC group; # P<0.05 compared with FT group. | |||
与HC组比较,FP组 、FT组和CC组小鼠血糖、血清总蛋白 和白蛋白水平均降低(P<0.05)。与CC组比较,FP组和FT组小鼠血糖、血清总蛋白和白蛋白 水平均增加(P<0.05),且FP组各项生化指标变化优于FT组(P<0.05)。见表3。
| (n=8,±s) | |||
| Group | Glucose [cB /(mmol·L-1)] | Total protein [ρB /(g·L-1)] | Albumin [ρB /(g·L-1)] |
| HC | 6.76±0.50 | 55.41±4.37 | 16.70±0.75 |
| CC | 3.42±0.31 * | 38.37±2.72 * | 12.40±0.94 * |
| FP | 5.53±0.49 * △# | 46.96±1.94 * △# | 15.16±0.53 * △# |
| FT | 4.32±0.45 * △ | 43.34±1.10 * △ | 13.96±0.25 * △ |
| * P<0.05 compared with HC group; △ P<0.05 compared with CC group; # P<0.05 compared with FT group. | |||
FP组、FT组和CC组小鼠腓肠肌质量及其肌纤维横切面积均低于HC组(P<0.05); FT组小鼠 腓肠肌质量及其肌纤维横切面积较CC组分别增长了25.43%和26.94%(P<0.05),FP组小鼠腓肠肌质量及其肌纤维横切面积较CC组分别增加了48.15%和56.19%(P<0.05),且FP组优于 FT组(P<0.05) 。见表4和图1(插页一)。
| (n=8,±s) | ||
| Group | Weight of GA(m/mg) | Myofiber cross-sectional area of GA (A/ μm2) |
| HC | 139.42±15.75 | 2384.56±138.92 |
| CC | 77.05±13.65 * | 1338.39±179.08 * |
| FP | 114.15±7.54 * △# | 2090.50±79.58 * △# |
| FT | 96.65±7.37 * △ | 1698.89±203.86 * △ |
| * P<0.05 compared with HC group; △ P<0.05 compared with CC group; # P<0.05 compared with FT group. | ||
与HC组比较,FP组、FT组和CC组小鼠腓肠肌组织中Mstn和Caspase-3 mRNA表达水平增加(P<0.05),而LncRNA-MALAT1mRNA表达水平降低(P<0.05);经FST干预后,FP组和FT组小鼠腓肠肌组织中Mstn和Caspase-3 mRNA表达水平均低于CC组(P<0.05),LncRNA-MALAT1mRNA表达水平均高于CC组(P<0.05),且FP组早期预防效果优于FT组(P<0.05)。FP组、FT组和CC组小鼠肿瘤组织LncRNA-MALAT1 mRNA表达水平无明显变化(P>0.05)。见表5。
| (n=3,±s) | ||||
| Group | Mstn | Caspase-3 | LncRNA-MALAT1 | |
| GA | Tumor | |||
| HC | 1.00±0.00 | 1.00±0.00 | 1.00±0.00 | - |
| CC | 5.26±0.49 * | 7.18±0.81 * | 0.32±0.07 * | 1.05±0.11 |
| FP | 1.91±0.28 * △# | 2.72±0.61 * △# | 0.76±0.14 * △# | 1.00±0.00 |
| FT | 3.11±0.47 * △ | 4.34±1.11 * △ | 0.55±0.12 * △ | 0.96±0.14 |
| “-”:No data. * P<0.05 compared with HC group; △ P<0.05 compared with CC group; # P<0.05 compared with FT group. | ||||
与HC组比较,FP组、FT组和CC组小鼠腓肠肌组织中Mstn和Caspase-3蛋白表达水平均升高(P<0.05);与CC组比较,FT组小鼠腓肠肌组织中Mstn和Caspase-3蛋白表达水平分别下降25.46%和35.51%(P<0.05),FP组小鼠腓肠肌组织中Mstn和Caspase-3蛋白表达水平分别下降51.96%和40.18%(P<0.05);且FP组小鼠腓肠肌组织中Mstn蛋白表达水平低于FT组(P<0.05) 。各组小鼠肿瘤组织中Caspase-3蛋白表达水平无明显变化(P>0.05)。见图2、3和表6。

|
| Lane 1:HC group;Lane 2:FP group;Lane 3:FT group;Lane 4:CC group. 图2 各组小鼠腓肠肌组织中Mstn和Caspase-3蛋白表达电泳图 Fig.2 Electrophoregram of Mstn and Caspase-3 protein expressions in GA tissue of mice in various groups |

|
| Lane 1:FP group;Lane 2:FT group;Lane 3:CC group. 图3 各组小鼠肿瘤组织中Caspase-3蛋白表达电泳图 Fig.3 Electrophoregram of Caspase-3 protein expressions in tumor tissue of mice in various groups |
| (n=3,±s) | |||
| Group | Mstn | Caspase-3 | |
| GA | Tumor | ||
| HC | 0.38±0.02 | 0.14±0.05 | - |
| CC | 1.24±0.14 * | 0.71±0.19 * | 0.47±0.05 |
| FP | 0.64±0.10 * △# | 0.43±0.11 * △ | 0.49±0.06 |
| FT | 0.95±0.09 * △ | 0.46±0.10 * △ | 0.55±0.05 |
| “-”:No data. * P<0.05 compared with HC group; △ P<0.05 compared with CC group; # P<0.05 compared with FT group. | |||
肿瘤恶病质是以短期内体质量下降为主要表现,同时以伴有食欲减退、全身衰竭及三大营养物质代谢紊乱为特征的临床综合征[1]。研究[2]表明:22%的恶性肿瘤患者死亡的直接原因为恶病质,而不是肿瘤本身。进行性骨骼肌组织减少是肿瘤恶病质发生发展的重要基础,能否缓解或逆转骨骼肌消耗成为预防和治疗肿瘤恶病质的关键。
Mstn是转换生长因子β(TGF-β)超家族的一员,其主要表达于骨骼肌中,对肌肉的生长发挥着负性调控作用。目前研究[3, 4, 5]显示:Mstn在肿瘤恶病质发生发展中发挥重要作用。抑制Mstn成为目前研究的热点[11]。FST是一种有效的Mstn抑制剂,能够阻断Mstn对骨骼肌的负性调控作用[9]。本研究结果显示:与CC组比较,经FST干预后FP组和FT组小鼠体质量和自发性活动量增加、腓肠肌质量及其肌纤维横切面面积均增加,营养状况得到改善,且Mstn mRNA和蛋白表达水平均明显降低。本文作者推测FST可通过抑制Mstn表达对肿瘤恶病质骨骼肌消耗起保护作用。
LncRNA-MALAT1是一种在哺乳动物多种组织中广泛表达的LncRNA,也是最早发现的与人类疾病相关的LncRNA[12]。LncRNA-MALAT1主要聚集于细胞核内核小斑结构中,通过招募SRSF1、SC35和 SRSF3 等多种剪接因子,介导mRNA前体加工过程中的可变剪接,从转录或转录后水平影响基因的表达[13, 14]。研究[15]显示:LncRNA-MALAT1能调控成肌细胞增殖与分化,在肌肉生发过程中发挥重要作用;Watts等[6]还发现:LncRNA-MALAT1在Mstn介导的肌肉消耗模型中表达显著下降,LncRNA-MALAT1表达异常可能与Mstn介导的骨骼肌消耗有关。LncRNA-MALAT1能够调控Caspase-3的表达[7],而Caspase-3在肿瘤恶病质骨骼肌降解过程中发挥着启动作用,其能水解相对分子质量大、结构复杂的肌蛋白为其他蛋白水解酶提供底物,还能够裂解特定的蛋白酶体亚基增加泛素蛋白酶体系统的活性,加速蛋白质降解,促进肿瘤恶病质发生发展[8]。
本研究结果显示:CC组小鼠由于肿瘤刺激使得Mstn表达增加引起LncRNA-MALAT1表达下降,Caspase-3表达升高,促进了骨骼肌降解,因此小鼠体质量下降,自发性活动量减少,腓肠肌质量及其肌纤维横切面积减少;FP组和FT组小鼠经FST干预后,随着Mstn表达下降,LncRNA-MALAT1表达增加,使得Caspase-3表达下降,骨骼肌消耗得到缓解,因此小鼠体质量增加,自发性活动量增多,营养状况得到改善,腓肠肌质量及其肌纤维横切面积增加。FST早期预防作用优于治疗作用,可能因为早期肿瘤负荷小,对机体的刺激作用较弱,Mstn表达少。早期运用FST抑制了肿瘤生长和Mstn表达,使得LncRNA-MALAT1表达水平升高,抑制了Caspase-3表达,因此骨骼肌降解减少。同时本研究还观察到FST并没对肿瘤组织中LncRNA-MALAT1及Caspase-3表达产生影响,可能原因是FST影响LncRNA-MALAT1及Caspase-3表达是通过抑制Mstn来实现的,而Mstn具有肌肉特异性[16],在肿瘤组织中不表达,故FST特异性促进骨骼肌组织中LncRNA-MALAT1 表达并抑制Caspase-3表达,对肿瘤患者是安全有效的。
综上所述,LncRNA-MALAT1在肿瘤恶病质骨骼肌中表达水平明显降低,FST缓解肿瘤恶病质小鼠骨骼肌消耗可能是通过降低Mstn表达,增加LncRNA-MALAT1表达,从而下调Caspase3表达来实现的。但Mstn调控LncRNA-MALAT1表达的具体机制以及LncRNA-MALAT1还能通过调控哪些靶基因来影响肿瘤恶病质肌肉消耗,尚需进一步研究证实。
| [1] | Fearon K,Strasser F,Anker SD,et al.Definition and classification of cancer cachexia:an international consensus[J].Lancet Oncol,2011,12(5):489-495. |
| [2] | Tisdale MJ. Cancer cachexia[J]. Anticancer Drugs,1993,4(2):115-125. |
| [3] | 薛 鹏,陶 钧,赵 晖,等.Myostatin基因在荷瘤裸鼠腓肠肌组织中的表达[J].上海交通大学学报:医学版,2006,26(10):1143-1146. |
| [4] | Gallot YS,Durieux AC,Castells J,et al. Myostatin gene inactivation prevents skeletal muscle wasting in cancer[J]. Cancer Res,2014,74(24):7344-7356. |
| [5] | Thissen JP,Loumaye A. Role of Activin A and Myostatin in cancer cachexia[J]. Ann Endocrinol (Paris),2013,74(2):79-81. |
| [6] | Watts R,Johnsen VL,Shearer J,et al. Myostatin-induced inhibition of the long noncoding RNA Malat1 is associated with decreased myogenesis[J]. Am J Physiol Cell Physiol,2013,304(10):C995-1001. |
| [7] | Guo F,Li Y,Liu Y,et al. Inhibition of metastasis-associated lung adenocarcinoma transcript 1 in CaSki human cervical cancer cells suppresses cell proliferation and invasion[J]. Acta Biochim Biophys Sin (Shanghai),2010,42(3):224-229. |
| [8] | Du J,Wang X,Miereles C,et al. Activation of caspase-3 is an initial step triggering accelerated muscle proteolysis in catabolic conditions[J]. J Clin Invest,2004,113(1):115-123. |
| [9] | Lee SJ,Lee YS,Zimmers TA,et al. Regulation of muscle mass by follistatin and activins[J]. Mol Endocrinol,2010,24(10):1998-2008. |
| [10] | Aulino P,Berardi E,Cardillo VM,et al. Molecular,cellular and physiological characterization of the cancer cachexia-inducing C26 colon carcinoma in mouse[J]. BMC Cancer,2010,10:363. |
| [11] | Han HQ,Mitch WE. Targeting the myostatin signaling pathway to treat muscle wasting diseases[J]. Curr Opin Support Palliat Care,2011,5(4):334-341. |
| [12] | Ji P,Diederichs S,Wang W,et al. MALAT-1,a novel noncoding RNA,and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer[J]. Oncogene,2003,22(39):8031-8041. |
| [13] | Tripathi V,Ellis JD,Shen Z,et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation[J]. Mol Cell,2010,39(6):925-938. |
| [14] | Änkö ML,Müller-McNicoll M,Brandl H,et al. The RNA-binding landscapes of two SR proteins reveal unique functions and binding to diverse RNA classes[J]. Genome Biol,2012,13(3):R17. |
| [15] | Han X,Yang F,Cao H,et al. Malat1 regulates serum response factor through miR-133 as a competing endogenous RNA in myogenesis[J]. FASEB J,2015,29(7):3054-3064. |
| [16] | Iizuka K,Machida T,Hirafuji M.Skeletal muscle is an endocrine organ[J]. J Pharmacol Sci,2014,125(2):125-131. |