 2022, Vol. 31
2022, Vol. 31



2. 内蒙古自治区不动产登记与国土资源储备交易中心, 内蒙古 呼和浩特 010010;
3. 中国地质科学院 地质研究所, 北京 100037
2. Real Estate Registration and Land Resources Reserve Trading Center of Inner Mongolia, Hohhot 010010, China;
3. Institute of Geology, Chinese Academy of Geological Sciences, Beijing 100037, China
热河生物群是分布于亚洲东部早白垩世中期的著名古生物化石群, 地质时代距今约120~131 Ma, 包括轮藻、苔藓类、蕨类、裸子植物、被子植物、双壳类、腹足类、甲壳类、蛛型类、昆虫, 以及无颌类、鱼类、两栖类、龟鳖类、有鳞类、离龙类、翼龙类、恐龙类鸟类和哺乳类等20个门类的动植物化石[1-2].其中包括轰动世界的多种长羽毛恐龙和古鸟类以及早期被子植物, 是20世纪末期新发现的一个世界级古生物化石宝库.
早期研究认为, 我国热河生物群主要分布于冀北、辽西等地区[3].内蒙古除赤峰南部地区外, 大部分地区没有热河生物群分布.直到20世纪80年代内蒙古地区开展专题区域地质研究, 才有系统研究热河生物群的报道.近年来随着古生物化石保护与调查评价工作不断深入, 在内蒙古更多的地区新发现了热河生物群化石分布区域和重要的化石产地(图 1).

|
图 1 内蒙古地区热河生物群化石产地分布图 Fig.1 Distribution of fossil sites of Jehol Biota in Inner Mongolia |
内蒙古地区白垩系分布广泛, 东自呼伦贝尔市海拉尔盆地、大兴安岭两侧, 西至阿拉善盟额济纳旗马鬃山地区均有出露, 以陆相盆地和山间盆地以及火山、火山碎屑沉积广泛发育为显著特征[4-5].在构造地层分区上, 研究区包括东部滨太平洋构造区的西沉降带鄂尔多斯盆地, 西隆起带北段的大兴安岭, 中沉降带的华北平原北缘的阴山-燕山沉降带, 向西延伸至阿拉善盟西端.
2 内蒙古热河生物群化石产地 2.1 燕山北段赤峰市南部 2.1.1 宁城县宁城县位于内蒙古自治区赤峰市东南部, 自东往西分别与辽宁省朝阳市建平县、凌源市, 河北省平泉县、承德县、隆化县接壤.宁城县中生代地层十分发育, 著名的道虎沟生物群赋存于中、上侏罗统九龙山组和髫髻山组[6].从上世纪90年代末期开始, 陆续在下白垩统义县组、九佛堂组中发现了丰富的热河生物群重要化石, 其中孟氏中生鳗及多种鱼类、鸟类、恐龙类化石具有重要科学意义, 还有虾类、蜥蜴、满洲鳄、满洲龟、恐龙类及丰富的古植物化石(图 2).化石产地包括土门、西三家、西五家、东五家、七家、小铜匠沟、柳条沟、朝阳山、王营子等地.

|
图 2 宁城县热河生物群部分代表化石 Fig.2 Representative fossils of Jehol Biota in Ningcheng County a—昆虫化石(insect fossil); b—蝾螈化石(salamander fossil); c—植物化石(plant fossil) |
目前在红山区境内已发现2处较有价值的化石产地, 含化石层位主要为下白垩统义县组和九佛堂组.其中1处位于红山区西南部文钟镇西水泉村麒麟山黑山沟一带, 产有双壳类、腹足类、昆虫类、叶肢介、鱼类、爬行类(龟、鳄、翼龙、鸟臀类恐龙化石等).
2.1.3 松山区主要分布于松山区初头郎镇薛家地至大庙镇一带, 含化石地层为下白垩统九佛堂组.薛家地蒙古图沟发现有鱼类、鸟类、节肢动物、软体动物和植物化石; 大庙一带发现过中生鳗、尾羽龙、鸟类等珍稀化石.具有重要研究价值和保护利用价值.
2.2 阴山北麓 2.2.1 巴彦淖尔市乌拉特后旗巴隆乌拉巴隆乌拉热河生物群化石产地位于乌拉特后旗西南巴隆乌拉一带的巴音戈壁组, 含恐龙、鱼类、植物化石等(图 3).2008年该地区发现了兽脚类、鸟臀类恐龙化石, 另外还有东方叶肢介、三尾拟蜉蝣等热河生物群典型化石.

|
图 3 乌拉特后旗巴隆乌拉热河生物群部分代表化石 Fig.3 Representative fossils of Jehol Biota in Balongwula, Urad Houqi a—鱼类化石(fish fossil); b—植物化石(plant fossil) |
内蒙古西部地区早期一直没有热河生物群的报道, 直到2008年, 在赛乌素镇北楚鲁庙下白垩统地层中, 首次发现热河生物群的重要分子: 球蚬、东方叶肢介、三尾拟蜉蝣、介形虫、松柏类植物化石、球果化石, 还有狼鳍鱼和十分完整的禽龙化石等(图 4).该发现已将热河生物群的分布向西推进了1000 km以上, 极大扩展了热河生物群的分布范围.

|
图 4 楚鲁庙热河生物群化石产地部分代表化石 Fig.4 Representative fossils of Jehol Biota in Chulumiao area a—叶肢介化石(conchostraca fossil); b—蜉蝣类化石(ephemera fossil) |
在固阳县下白垩统李三沟组和固阳组中, 含有丰富的淡水湖泊生物群化石, 如双壳类、腹足类、介形类等无脊椎动物化石和狼鳍鱼化石.此外, 还发现有较完整的鹦鹉嘴龙化石.
2.3 鄂尔多斯盆地鄂尔多斯地区目前发现的热河生物群化石主要分布于鄂托克旗, 赋存在下白垩统上部泾川组灰色砂质泥岩中, 已发现的化石有狼鳍鱼、弓鳍鱼、多齿始马来鳄及新鳄形类、伊克昭龙、成吉思汗鄂托克鸟、查布华夏鸟等古脊椎动物化石(图 5).在查布地区周边相关的层位还发现有龟鳖类、翼龙类以及丰富的恐龙类等[7]; 查布地区无脊椎动物化石有介形虫、昆虫类、双壳类、腹足类等[8].值得一提的还有, 在查布地区早白垩世地层中还保存有数以万计精美的恐龙足迹和鸟类足迹化石[9-10].

|
图 5 鄂尔多斯盆地热河生物群化石产地部分代表化石 Fig.5 Representative fossils of Jehol Biota in Ordos Basin a—鱼类化石(fish fossil); b—鸟类足迹化石(bird footprint fossil); c—恐龙足迹化石(dinosaur footprint fossil) |
2014年在莫力达瓦旗发现了重要的热河生物群化石产地, 主要分布在莫力达瓦旗西南部的宝山镇下白垩统龙江组中.含化石岩层为黄灰色的凝灰质(粉砂质)泥页岩, 其中含有种类多样的古生物化石, 化石数量最多的为叶肢介化石, 此外还有昆虫类、鱼类(中生鳗、狼鳍鱼等)、两栖类(蝾螈类、蛙类)、龟鳖类、鸟类化石以及大型爬行动物骨骼化石, 还见有较多植物化石[11-16](图 6).从已有化石组合面貌看, 莫力达瓦生物群与辽西热河生物群具有非常高的相似性, 某种程度上其化石种类更加丰富多样.这是内蒙古地区首次发现内容如此丰富的早白垩世热河生物群, 对于热河生物群的古地理分布、生物群的面貌及演化特征研究具有十分重大的科研价值, 被评定为国家级重点保护古生物化石产地.

|
图 6 莫力达瓦达斡尔族自治旗热河生物群部分代表化石 Fig.6 Representative fossils of Jehol Biota in Daur Autonomous Banner of Morin Dawa a—蛙类化石(frog fossil); b—植物化石(plant fossil) |
值得一提的还有这里的两栖类化石数量多、密度大, 已发现在一块约1 m2的石板上有9个蛙化石和1个蝾螈化石共生, 这在我国绝无仅有.此外, 目前我国已发现的热河生物群化石产地中, 蛙类和蝾螈化石共生的情况极为罕见.
2.4.2 呼伦贝尔市阿荣旗在阿荣旗亚东镇和霍尔奇镇那克塔等地也发现有丰富的叶肢介、昆虫类和狼鳍鱼等热河生物化石群.
3 古气候与古环境讨论 3.1 燕山北段赤峰地区大地构造位置上, 赤峰地区位于华北板块与天山-兴蒙造山带两大构造单元拼合带北缘, 其构造形迹显示出受两大构造单元强烈影响的过渡带特征.在中生代燕山期, 北部鄂霍次克洋洋壳消亡, 东部古太平洋位置上伊佐奈歧-库拉板块北西向的挤压, 形成复合应力场, 受这种复合应力作用的影响, 形成了该区特定的构造格架, 即纬向构造与北北东向构造交替穿插出现。从整体上看, 赤峰盆地是一个凹凸相间、具多个次凹的复式凹陷, 属残留型盆地范畴.
赤峰地区中生界下白垩统义县组一段的岩性为灰色、灰黄色、灰黄绿色砂岩和泥质页岩, 显示较稳定沉积环境, 应为陆相湖泊[17].上覆浅紫红色火山角砾岩、流纹岩, 为陆地火山喷发溢流相环境(表 1).任东等人通过研究该地区昆虫化石, 发现绝大多数昆虫化石为水生的种类, 昆虫群的总体面貌与在蒙古国和俄罗斯贝加尔以及我国冀北、辽西、吉林等地区广泛分布的热河昆虫群基本一致, 反映当时该地与以上地区通过共同的水系沟通.根据特殊昆虫种属判断本地水流较缓、地形起伏不大, 应为湖泊或湖沼环境[18].赤峰地区热河生物群发现大量鱼类、两栖类和水生爬行类(满洲鳄和满洲龟)等脊椎动物化石, 及双壳类、腹足类和叶肢介等无脊椎动物化石, 也证明当时水系发育, 河流湖泊分布广泛.大量植物和孢粉化石进一步证明当时的古气候以温暖湿润的温带气候为主.
|
|
表 1 赤峰地区与周边地区古生物分布地层对比 Table 1 Stratigraphic correlation of paleontological distribution between Chifeng region and surrounding areas |
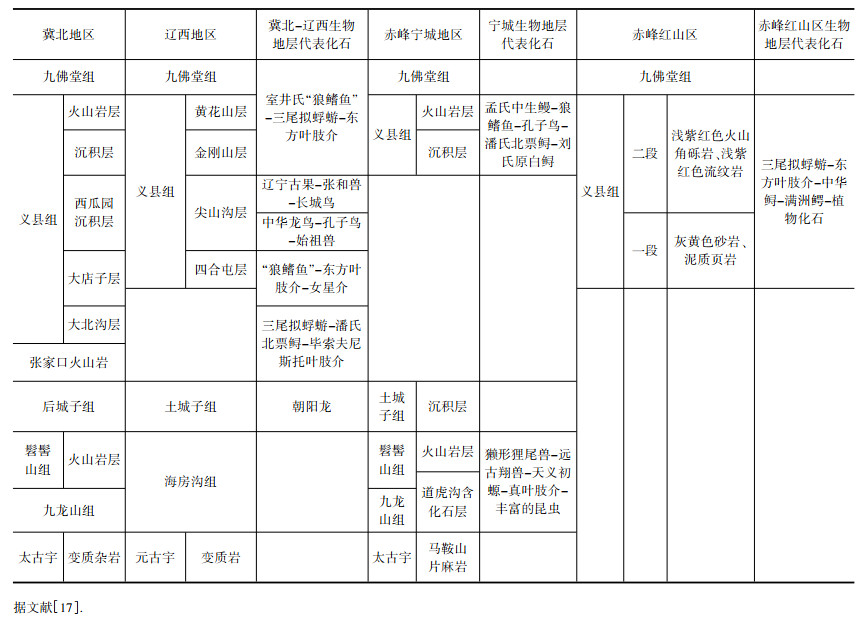
巴音戈壁组, 由宁夏区调队1980年命名, 命名剖面位于内蒙古乌拉特后旗乌拉山, 下部为灰白色或黄褐色砾岩、钙质长石细砂岩夹砂砾岩、页岩; 中部为灰黑色粉砂质灰岩; 上部为紫灰色或灰绿或蓝灰或灰黑色页岩、油页岩夹泥质粉砂岩, 显示出由冲洪积相-河流相逐渐转变为湖沼相、滨-浅湖相.根据岩性颜色特点巴音戈壁组分为下段的红色碎屑岩系和上段的灰色碎屑岩系, 分别反映沉积时的气候干旱炎热(氧化环境)和温暖潮湿(还原环境).
已有研究认为早白垩世华北北缘处于亚热带半干旱气候区的最南缘, 相对于松辽盆地较干燥而温暖[19].而据硅化木的发现显示银根-额济纳旗盆地早白垩世植被茂盛、气候温湿[4].孢粉化石组合则显示巴音戈壁组二段应为属早白垩世中晚期, 气候总体呈现温暖半干旱特征[20].再次证实内蒙古西部阿拉善一带在白垩纪早期经历了由干旱至温湿的气候转变.
3.3 鄂尔多斯盆地侏罗纪末期-早白垩世, 在燕山运动作用下, 鄂尔多斯盆地以南北向为主轴方向, 形成东高西低的非均衡不对称盆地, 西岸陡倾、东部和南部相对平缓, 盆地内基底台拗相间、起伏不平[21].这种盆地结构形态和古地理面貌直接影响沉积特征、岩相发育及展布规律.在盆地东翼, 由于坡降小, 地形开阔, 形成的洛河组厚层砂岩产状和厚度稳定, 相带宽, 结构均匀.而在盆地西翼, 地形陡, 坡降大, 从洛河组到泾川组, 河流与冲积扇沉积厚度变化大, 一般为100~800 m, 颗粒粗, 岩性组分复杂.
内蒙古鄂尔多斯地区的鄂托克旗和杭锦旗出露有较大面积的早白垩世罗汉洞组和泾川组, 岩性以紫红色和灰色砂岩、含砾粗砂岩为主及少量页岩.显示应为干旱-半干旱陆内河湖相沉积环境.
3.4 大兴安岭岭东地区岭东地区地质构造背景复杂, 先后经历了古亚洲洋闭合及滨太平洋构造运动的作用, 存在华力西和燕山两期岩浆活动.区内古生代地层出露较少, 见少量晚石炭世火山岩及中生代火山岩地层大面积出露, 按形成时代由老至新有下白垩统龙江组、光华组、甘河组、孤山镇组, 为一套湖盆相火山-沉积地层.秦涛等通过对龙江组孢粉化石属种的分析, 结合地质、古植物及孢粉化石的定量分析, 认为龙江组植被类型反映亚热带气候, 指示温暖潮湿的环境[22-23].
整体上, 内蒙古地区的热河生物群由于地域差别, 东、西两部的化石组成有较大的不同.东部地区主要分布在赤峰市南部宁城地区, 地质构造上属于辽西大型沉积盆地的北部, 其生物群组成与辽西基本一致.而内蒙古其他地区, 主要包括大兴安岭、西部阿拉善盟、巴彦淖尔市等地, 其热河生物群与辽西的热河生物群有较大的区别, 特别是恐龙生物群, 除了禽龙外, 地方色彩十分明显.巴彦淖尔市的热河生物群出现了乌拉特半爪龙、单爪龙等地方特有的恐龙属种[24].无脊椎动物中的叶肢介, 是以尼斯托叶肢介为主, 而不是内蒙古东部的东方叶肢介为主.狼鳍鱼数量较少, 代之的是中华弓鳍鱼.植物化石数量少, 品种单调.
上述特征均反映了地域差别而导致的生物群组成分子的差别.说明中国的热河生物群自辽西向西逐渐衰弱的趋势.这可能是由于东部的火山活动比较强烈, 形成的火山灰的沉积物较为丰富, 从而保存了众多鱼类、鸟类、爬行类、原始哺乳动物, 以及大量无脊椎动物化石和植物化石, 从而形成了种类丰富、数量众多的热河生物化石群.而西部地区, 缺乏东部地区的沉积环境, 化石保存情况差, 加之生物种类单调, 导致发现的化石种类较少, 形成了明显的地域差异.
4 国内外热河生物群对比讨论目前狭义的热河生物群分布区域包括辽宁西部、河北北部和内蒙古东南部, 而广义的热河生物群分布区域还包括中国新疆、甘肃、内蒙古中西部及大兴安岭、陕西、吉林、山东、浙江等地区, 及蒙古国、哈萨克斯坦、俄罗斯、日本和朝鲜半岛[1].根据热河生物群生物面貌组合特征, 目前已识别出3个演化阶段: 早期(冀北期)、中期(热河早期)和晚期(热河晚期)[25].内蒙古自治区内热河生物群化石产地既包括狭义的核心分布区(赤峰地区), 也有广义分布区(阴山北麓、鄂尔多斯盆地和大兴安岭地区), 所有化石产地均发现狼鳍鱼、三尾拟蜉蝣和东方叶肢介, 属于热河生物群中期演化阶段.热河生物群在河北省内主要分布于承德地区, 赋存层位包括大北沟组和花吉营组(义县组), 化石属种比较单调, 但包含许多热河生物群早期分子, 如原始的反鸟类(丰宁原羽鸟)和今鸟型类(弥曼始今鸟), 代表热河生物群早期发展阶段.辽宁西部(朝阳、锦州、葫芦岛)是热河生物群分布最核心地区, 化石主要赋存于义县组和九佛堂组.由于缺失可与冀北地区大北沟组对比的地层, 单从生物群组合面貌分析, 辽西地区热河生物群在属种丰度、化石数量和保存状态等方面遥遥领先于世界其他地区, 既包括早期被子植物(辽宁古果)、早期有胎盘类哺乳动物(攀援始祖兽)和早期原始鸟类(热河鸟类)等, 也包括进步的鸟类(胡氏旅鸟), 代表了热河生物群早期和中期发展阶段.热河生物群在甘肃省内主要分布于玉门地区, 化石赋存于下沟组(< 120 Ma), 生物类群比较单调, 根据发现的鸟类化石主要包括比较进步的今鸟型类(甘肃鸟), 缺少比较原始的鸟类(基干鸟类), 代表了热河生物群晚期发展阶段.国内其他地区(陕西、吉林、山东、浙江等)发现的热河生物群化石数量较少, 属种分异度很低, 分布面积小, 研究程度低, 很难进行深入的横向对比.国外发现的热河生物群化石属种相对比较单调, 目前仅在日本福井县发现少量恐龙和鸟类化石, 俄罗斯西伯利亚地区发现孔子鸟类的部分骨骼化石, 其他主要是无脊椎动物、昆虫和鱼类化石.由于国外的化石资料较少, 地层对比研究也存在争论, 无法准确进行对比讨论.
热河生物群地理分布的广泛性及包含化石门类的多样性, 均为白垩纪古气候和古环境研究提供了重要信息, 其中植物对气候变化最为敏感.热河生物群早期孢粉植被以针叶林为主, 逐渐转变为以灌木和草本植物为主, 说明气温逐渐升高.但许多长羽毛恐龙的出现(尤其是早期原始羽毛不具飞行功能, 主要用来保持体温), 说明热河期曾存在气候相对寒冷的短暂时期, 但寒冷期的持续时间及影响范围有待后期深入研究.即使在热河生物群核心分布区, 不同化石产地或同一化石产地不同阶段也存在不同的小气候环境.目前关于热河生物群的研究多侧重于单一类群(或门类)的系统演化或单一领域(古生物学、地球化学、年代学等)的研究.全面而系统地研究整个热河生物群起源、演化、绝灭和该地质时期的气候变化、地质环境变迁及其相互作用仍处于起步阶段.
5 结论基于对内蒙古自治区东、西部白垩纪热河生物群化石分布特征及地层对比研究, 可以得出以下结论:
(1) 热河生物群在内蒙古自治区内主要分布于东部区呼伦贝尔市莫力达瓦达斡尔族自治旗和阿荣旗的龙江组, 赤峰市东南部义县组和九佛堂组; 中部区鄂尔多斯盆地鄂托克旗查布地区的泾川组; 西部乌拉特后旗巴隆乌拉的巴音戈壁组, 固阳盆地固阳县的李三沟组和固阳组.
(2) 内蒙古东、西部白垩纪热河生物群对比研究发现, 东部区古生物化石极其丰富, 生物群组成与辽西基本一致, 脊椎动物主要包括无颌类、硬骨鱼类、两栖类、爬行类、鸟类和哺乳动物等.而西部古生物化石相对较少, 生物群组成也存在较大区别, 出现了具有地方特色的禽龙类和阿尔瓦雷斯龙类等特殊种属.
(3) 内蒙古东部白垩系主要以火山岩、凝灰岩为主.受燕山期构造影响明显, 火山活动极其频繁; 古气候方面主要为亚热带温热潮湿的环境, 为内陆湖相或湖沼相沉积.而西部地区白垩系主要为黄褐色、紫红色的碎屑沉积岩, 火山岩整体较少, 显示出干旱-半干旱的气候为主, 局部可见较湿润气候, 判断为陆内河流相或河湖相.
(4) 内蒙古白垩纪时期沉积古地理、古气候与古生态环境演化与热河生物群辐射演化具有显著的制约和协同关系.内蒙古白垩纪热河生物群各个重要门类的发现和研究为认识早白垩世全球陆地生态系统生物多样性的构成和辐射演化提供了重要的化石证据.
| [1] |
周忠和, 王原. 热河生物群脊椎动物生物多样性的分析以及与其他动物群的比较[J]. 中国科学: 地球科学, 2010, 53(9): 1250-1265. Zhou Z H, Wang Y. Vertebrate diversity of the Jehol Biota as compared with other lagerstätten[J]. Science China Earth Sciences, 2010, 53(12): 1894-1907. |
| [2] |
徐星, 周忠和, 王原, 等. 热河生物群研究的回顾与展望[J]. 中国科学: 地球科学, 2020, 63(10): 1491-1511. Xu X, Zhou Z H, Wang Y, et al. Study on the Jehol Biota: Recent advances and future prospects[J]. Science China Earth Sciences, 2020, 63(6): 757-773. |
| [3] |
顾知微. 中国侏罗系和白垩系[J]. 科学记录新辑, 1960, 4(4): 202-212. Gu Z W. Jurassic and cretaceous in China[J]. New Scientific Records, 1960, 4(4): 202-212. |
| [4] |
张明震, 戴霜, 陈世强, 等. 内蒙古银根-额济纳旗盆地白垩系中硅化木化石及其沉积环境[J]. 地质学报, 2013, 87(8): 1059-1066. Zhang M Z, Dai S, Chen S Q, et al. Early Cretaceous silicied woods from the Yingen-Ejinaqi Basin, Inner Mongolia and their sedimentary environment[J]. Acta Geologica Sinica, 2013, 87(8): 1059-1066. DOI:10.3969/j.issn.0001-5717.2013.08.002 |
| [5] |
吴仁贵, 周万蓬, 刘平华, 等. 关于内蒙古巴音戈壁盆地早白垩世地层的讨论[J]. 地层学杂志, 2009, 33(1): 87-90. Wu R G, Zhou W P, Liu P H, et al. Discussion on the Early Cretaceous in the Bayingebi Basin[J]. Journal of Stratigraphy, 2009, 33(1): 87-90. DOI:10.3969/j.issn.0253-4959.2009.01.011 |
| [6] |
谭京晶, 任东. 内蒙古宁城中侏罗世九龙山组昆虫群落生态的初步研究[J]. 动物分类学报, 2002, 27(3): 428-434. Tan J J, Ren D. Palaeoecology of insect community from Middle Jurassic Jiulongshan Formation in Ningcheng County, Inner Mongolia, China[J]. Acta Zootaxonomica Sinica, 2002, 27(3): 428-434. DOI:10.3969/j.issn.1000-0739.2002.03.003 |
| [7] |
姬书安, 张笠夫, 卢立伍, 等. 内蒙古鄂尔多斯地区早白垩世翼龙类化石的发现[C] //中国古生物学会第28届学术年会论文摘要集. 沈阳: 中国古生物学会, 2015: 165-166. Ji S A, Zhang L F, Lu L W, et al. The discovery of Early Cretaceous pterosaur fossils from the Ordos region, Inner Mongolia[C] //Proceedings of the 28th Annual Conference of Palaeontological Society of China. Shenyang: Palaeontological Society of China, 2015: 165-166. |
| [8] |
崔新省, 刘本培. 鄂尔多斯盆地西南部华亭-陇县志丹群泾川组的双壳类[J]. 古生物学报, 1994, 33(4): 429-439. Cui X S, Liu B P. Bivalves from Jingchuan Formation of Zhidan Group between Huating and Longxian, southwestern Ordos Basin[J]. Acta Palaeontologica Sinica, 1994, 33(4): 429-439. |
| [9] |
王宝鹏, 李建军, 白志强, 等. 内蒙古鄂托克旗查布地区恐爪龙类足迹的发现及其意义[J]. 北京大学学报(自然科学版), 2017, 53(1): 81-90. Wang B P, Li J J, Bai Z Q, et al. The first discovery of deinonychosaurian tracks from Lower Cretaceous of Chabu, Otog Qi, Inner Mongolia and its significance[J]. Acta Scientiarum Naturalium Universitatis Pekinensis, 2017, 53(1): 81-90. |
| [10] |
李建军, 巴特尔, 张维虹, 等. 内蒙古查布地区下白垩统巨齿龙足印化石[J]. 古生物学报, 2006, 45(2): 221-234. Li J J, Bater, Zhang W H, et al. A new type of dinosaur tracks from Lower Cretaceous of Chabu, Otog Qi, Inner Mongolia[J]. Acta Palaeontologica Sinica, 2006, 45(2): 221-234. DOI:10.3969/j.issn.0001-6616.2006.02.006 |
| [11] |
王龙, 程晓冬, 李伟, 等. 中国内蒙古大兴安岭地区下白垩统光华组白垩逍遥蛛一新种[J]. 中国地质, 2017, 44(4): 818-819. Wang L, Cheng X D, Li W, et al. A new species of Cretadromus from the Lower Cretaceous Guanghua Formation in the Da Hinggan Mountains, Inner Mongolia[J]. Geology in China, 2017, 44(4): 818-819. |
| [12] |
Jia J, Gao K Q. A new hynobiid-like salamander (Amphibia, Urodela) from Inner Mongolia, China, provides a rare case study of developmental features in an Early Cretaceous fossil urodele[J]. Peer J, 2016, 4: e2499. DOI:10.7717/peerj.2499 |
| [13] |
Gao K Q, Chen J Y. A new crown-group frog (Amphibia: Anura) from the Early Cretaceous of northeastern Inner Mongolia, China[J]. American Museum Novitates, 2017, 3876: 1-39. DOI:10.1206/3876.1 |
| [14] |
Wang X R, Wang L, Ji Q. First discovered diversified fossil locality of the Jehol Biota in the Greater Khingan Mountains, Inner Mongolia[J]. Acta Geologica Sinica (English Edition), 2017, 91(5): 1906-1907. DOI:10.1111/1755-6724.13420 |
| [15] |
Xing L D, Niu K C, Evans S E. Inter-amphibian predation in the Early Cretaceous of China[J]. Scientific Reports, 2019, 9(1): 7751. DOI:10.1038/s41598-019-44247-7 |
| [16] |
Wang X R, Cau A, Kundrát M, et al. A new advanced ornithuromorph bird from Inner Mongolia documents the northernmost geographic distribution of the Jehol paleornithofauna in China[J]. Historical Biology, 2021, 33(9): 1705-1717. DOI:10.1080/08912963.2020.1731805 |
| [17] |
王政恺. 内蒙古赤峰市红山区麒麟山中生代热河生物群化石发现及意义[D]. 北京: 中国地质大学, 2011. Wang Z K. Discovery and significance of Mesozoic Jehol Biota in Qilin Hill, Hongshan District, Chifeng City, Inner Mongolia[D]. Beijing: China University of Geosciences, 2011. |
| [18] |
任东, 朱会忠, 陆有泉. 内蒙古赤峰早白垩世昆虫化石的新发现[J]. 地球学报, 1995, 17(4): 432-439. Ren D, Zhu H Z, Lu Y Q. New discovery of Early Cretaceous fossil insects from Chifeng City, Inner Mongolia[J]. Acta Geosicientia Sinica, 1995, 17(4): 432-439. |
| [19] |
付国斌, 李罡, 任玉光, 等. 内蒙古呼伦陶勒盖下白垩统巴音戈壁组的叶肢介化石[J]. 古生物学报, 2007, 46(2): 244-248. Fu G B, Li G, Ren Y G, et al. Early Cretaceous conchostracans from the Bayingebi Formation of Inner Mongolia, China[J]. Acta Palaeontologica Sinica, 2007, 46(2): 244-248. |
| [20] |
杨友运. 鄂尔多斯盆地白垩系沉积建造[J]. 石油与天然气地质, 2006, 27(2): 167-172. Yang Y Y. Cretaceous sedimentary formation in Ordos Basin[J]. Oil & Gas Geology, 2006, 27(2): 167-172. |
| [21] |
任文秀, 赵海钧, 胡永兴, 等. 鄂尔多斯盆地西南缘早白垩世地层孢粉组合及地层意义[J]. 地质论评, 2019, 65(S1): 129-131. Ren W X, Zhao H J, Hu Y X, et al. Early Cretaceous sporopollen assemblage and stratigraphic significance in southwestern margin of Ordos Basin[J]. Geological Review, 2019, 65(S1): 129-131. |
| [22] |
秦涛, 李林川, 钱程, 等. 内蒙古扎兰屯地区龙江组孢粉组合及其地质意义[J]. 地质通报, 2018, 37(9): 1571-1578. Qin T, Li L C, Qian C, et al. The palynological assemblage of the Longjiang Formation in Zhalantun area, Inner Mongolia, and its geological implications[J]. Geological Bulletin of China, 2018, 37(9): 1571-1578. |
| [23] |
钱程, 汪岩, 秦涛, 等. 内蒙古扎兰屯地区晚三叠世韧性剪切变形的激光40Ar/39Ar年龄证据[J]. 中国地质, 2022, 49(1): 346-347. Qian C, Wang Y, Qin T, et al. Evidence from muscovite laser 40Ar/39Ar age for Late Triassic ductile shear deformation in Zhalantun area, Inner Mongolia[J]. Geology in China, 2022, 49(1): 346-347. |
| [24] |
徐星. 单爪龙的侏罗纪祖先[J]. 化石, 2014(3): 45-55. Xu X. The ancestors of mononykus in Jurassic Period[J]. Fossils, 2014(3): 45-55. |
| [25] |
万晓樵, 李罡, 黄清华, 等. 中国白垩纪陆相阶的划分与对比[J]. 地层学杂志, 2013, 37(4): 457-471. Wan X Q, Li G, Huang Q H, et al. Division and correlation of terrestrial Cretaceous stages in China[J]. Journal of Stratigraphy, 2013, 37(4): 457-471. |