 2022, Vol. 31
2022, Vol. 31


2. 沈阳师范大学 古生物学院, 辽宁 沈阳 110034;
3. 东北亚国际地学研究与教学中心, 吉林 长春 130026
2. College of Paleontology, Shenyang Normal University, Shenyang 110034, China;
3. International Center for Research and Education of Geosciences in Northeast Asia, Changchun 130026, China
白垩纪(145~66 Ma)是中生代最后一个纪,也是地球生物/地质演化最复杂与最精彩的时段之一. 此间的“温室”气候、有花植物的出现、恐龙从鼎盛到灭绝、石油与煤的大量形成、强烈的火山活动等特征,都是这一时期的“亮点”.
中国东北地区东部(主要指黑龙江省东部和吉林省中东部)白垩纪陆相地层十分发育,富含动、植物化石. 这一地区的白垩纪地层古生物研究已有近百年历史,大体上经历了早期认识(1920—1960年代)、区调及专题研究(1970—1980年代)、新发现与深入研究(1990—2000年代)以及多学科交叉研究与新突破(21世纪起)等4个阶段. 然而,由于这一地区地层出露较为零星,动、植物化石的分类及时代等又长期存在争论,加之缺少同位素年代学研究等,有关白垩纪地层及化石有许多问题需要解决.
近年来,随着各门类化石研究的不断深入,特别是同位素年代学的研究水平迅速提升与广泛参与,东北地区东部白垩纪生物地层学研究取得了诸多重要成果,部分成果已跻身于国际前列. 本文仅就这一地区的有关白垩纪植物群及其地层研究的一些进展做简要介绍,并对晚白垩世该地区植物群与以恐龙为代表的动物群协同演化进行初步探讨.
1 早白垩世植物群及地层研究进展 1.1 滴道组植物化石及鸡西盆地早白垩世最早期海侵黑龙江东北部饶河—绥滨地区的早白垩世海相地层是我国除西藏外唯一留存的早白垩世早期(贝里阿斯—瓦兰今期,即Berriasian-Valanginian)海相地层记录. 1982年黑龙江省第一区调大队在饶河东安镇首次建立了东安镇组(J3—K1)[1]. 1985年沙金庚等首先描述了这里的海相双壳类雏蛤(Buchia)化石[2]. 1986年笔者率课题组在东安镇组典型剖面首次划分了上、下2个亚组和4个化石带(B1—B4),以B2/B3交界作为侏罗—白垩系(J—K)界线:该界线之下以Buchia russiensis为代表,属提塘阶(Tithonian,J3);其上以B. unschenesis,B. volgensis及B. okensis等为代表,属贝里阿斯—瓦兰今阶(Berriasian-Valanginian,K1)[3-4](图 1a,c—e). 绥滨地区相应的J—K界线附近海相地层称东荣组,由钻孔揭露,也分为上、下两部分[4-6](图 1b). 饶河—绥滨地区的上述发现为研究黑龙江东部早白垩世早期海陆分布等打下了坚实基础. 本世纪初,程金辉等[7]及祝幼华等[8]在鸡西盆地滴道组发现早白垩世早期(Berriasian-Valanginian)海相沟鞭藻化石,为认识鸡西盆地早白垩世早期的海侵提出了新的思考.

|
图 1 黑龙江饶河—绥滨早白垩世海相地层及东安镇海相J3—K1剖面 Fig.1 Early Cretaceous marine strata in Raohe-Suibin area and marine J3—K1 section in Dong'anzhen a—中国早白垩世地层分布图(蓝色示海相,粉色示陆相,据文献[6]修改)(distribution of Lower Cretaceous strata in China,blue for marine facies,pink for continental facies,modified from Reference[6]);b—笔者率课题组在钻井旁查看绥滨组岩心(the author’s research team checking the borehole cores in Suibin);c—东安镇组上亚组剖面(K1瓦兰今阶)(section of Upper Dong’anzhen fm.,Valanginian);d—饶河东安镇海相J3—K1剖面图(marine J3—K1 section in Dong’anzhen);e—东安镇组海相Buchia化石(marine bivalves Buchia from Dong’anzhen fm.):1—Buchia russiensis(东安镇组下亚组,J3提塘阶)(Lower Dong’anzhen fm.,Tithonian),2—B. unschensis,3—B. okensis,4—B. volgensis (2,3,4为东安镇组上亚组,K1贝里阿斯阶—瓦兰今阶;c,d,e据文献[3])(2,3,4 from Upper Dong’anzhen fm.,Berriasian-Valanginian;c,d,e:from Reference[3]) |
鸡西盆地滴道组的植物化石已有多年研究历史:郑少林与张武曾在该组发现18属25种植物化石[9];此后,笔者与郑少林等又对滴道组植物化石进一步采集和研究[10]. 前不久,在冯玉辉研究滴道组火山岩期间,笔者对在鸡西荣丰三队滴道组典型剖面采得的松柏类化石——莱恩苏铁杉(Podozamites reinii Geyler)做了进一步时代对比研究,结果表明:该种最早发现于日本石川县桑岛市白岩村(Shiramine)的桑岛组(Kuwajima Formation)[11],该组时代一直被认为是Berriasian-Valanginian期[12],后发现该组也有凝灰岩同位素测年(130.2 Ma)的报道[13-14]. 经笔者委托日本同行代查,日本国立科学博物馆确收藏有Podozamites reinii标本(登记号N200600236),产自Valanginian期地层(H. Takimoto函告,2022). 由此说明,莱恩苏铁杉(P. reinii)至少最早有出现在Valanginian期的记录. 这一时代认识为冯玉辉等首次测得滴道组131.5±1.4 Ma年龄[15],认为整个滴道组时代可能为早白垩世瓦兰今—欧特里夫期(Valanginian-Hauterivian)提供了植物化石的辅证. 结合程金辉等[6]及祝幼华等[7]在滴道组发现的以Lagenorhytis granorugosus-Vesperopsis didaoensis组合为代表的海相沟鞭藻化石,冯玉辉等认为Valanginian期可能海水已侵入鸡西盆地(以滴道组海相层位为代表),这应是鸡西盆地最早的海侵记录[15]. 另值得提及的是滴道组两个颇具特征的植物分类群,滴道拟蕨(Pteriopsis didaoensis Zheng et Zhang)和莱恩苏铁杉(Podozamites reinii Geyler),在滴道组上部的城子河组并未发现过.
1.2 鸡西早期被子植物新知黑龙江省鸡西地区城子河组早期被子植物化石(以下简称“鸡西早期被子植物”)是黑龙江省东部地区迄今最早的被子植物,也是研究中国东北地区早期被子植物演化发展历程的重要“节点”之一. 鸡西早期被子植物的植物学特征、时代等也关系到我国与全球早期被子植物的对比,一直备受关注[10, 16-20].
1990—1992年由笔者率课题组首次发现的早白垩世鸡西早期被子植物为揭示我国东北地区东部早期被子植物的发生与演化提供了重要资料(图 2a). 鸡西早期被子植物至少包括亚洲叶(Asiatifolium)、鸡西叶(Jixia)、沈括叶(Shernkuoa)、郑氏叶(Zhengia)及星学花序(Xingxueiana)等9个分类群[16-18](图 2b). 以往笔者等认为其时代大体为巴雷姆期或巴雷姆晚期—阿普特早期[10, 16-19, 21-22]. 但近年来通过鸡西早期被子植物在俄罗斯南滨海地区早白垩世偏晚些时代的地层中被发现[20, 23]以及相关发现[24],城子河组相关动物化石(如Aucellina等)[25-27]及同位素测年所提供的参考(图 2c),兼考虑城子河组下部海相层(第12层)的时代[28],目前似可考虑鸡西城子河组全组的时代可能为巴雷姆晚期—阿普特早期(late Barremian-early Aptian)(表 1);而城子河组的被子植物化石(即鸡西早期被子植物,位于该组中上部)的时代可能为阿普特早期(early Aptian).

|
图 2 黑龙江鸡西植物群的早期被子植物及其地层 Fig.2 Early angiosperm from Jixi flora in Heilongjiang Province and its strata a—鸡西城子河早白垩世城子河组剖面(李春田摄,2006)(profile of Lower Cretaceous Chengzihe fm. in Jixi);b—城子河组早期被子植物化石(early angiosperm fossil of Chengzihe fm.):1,2,3—优美亚洲叶(Asiatifolium elegans),4—羽叶鸡西叶(Jixia pinnatipartita),5,8—城子河鸡西叶(J. chengzihensis),6—奇脉沈括叶(Shenkuoa caloneura),7,12—鸡西叶(未定种)(J. sp.),9,10,11—星学花序(Xinxueiana),9—花序(inflorescence),10,11—无沟花粉(inapertural pollen)(1~11据文献[10, 16-18]),12为俄罗斯南滨海标本,E. Volynets提供)(Figs. 1—11 from References[10, 16-18],12 from South Primorye of Russia,by courtesy of E. Volynets);c—鸡西盆地及邻区早白垩世地层分布及其火山岩年龄(据文献[15])(distribution of Lower Cretaceous strata in Jixi Basin and adjacent areas with their volcanic rock ages,from Reference[15]));d~h—俄罗斯南滨海地区发现鸡西早期被子植物(据文献[23])(early angiosperm of Jixi flora found in South Primorye of Russia,from Reference[23]):d,e—俄罗斯南滨海波手-卡门剖面及化石产出层位(Bolshoy Kamen section and fossil-bearing horizon in South Primorye of Russia),f—羽叶鸡西叶(Jixia pinnatipartita),g,h—优美亚洲叶(Asiatifolium elegans) |
|
|
表 1 中国东北地区东部白垩纪地层划分与对比简表 Table 1 Division and correlation of the Lower Cretaceous strata in eastern Northeast China |
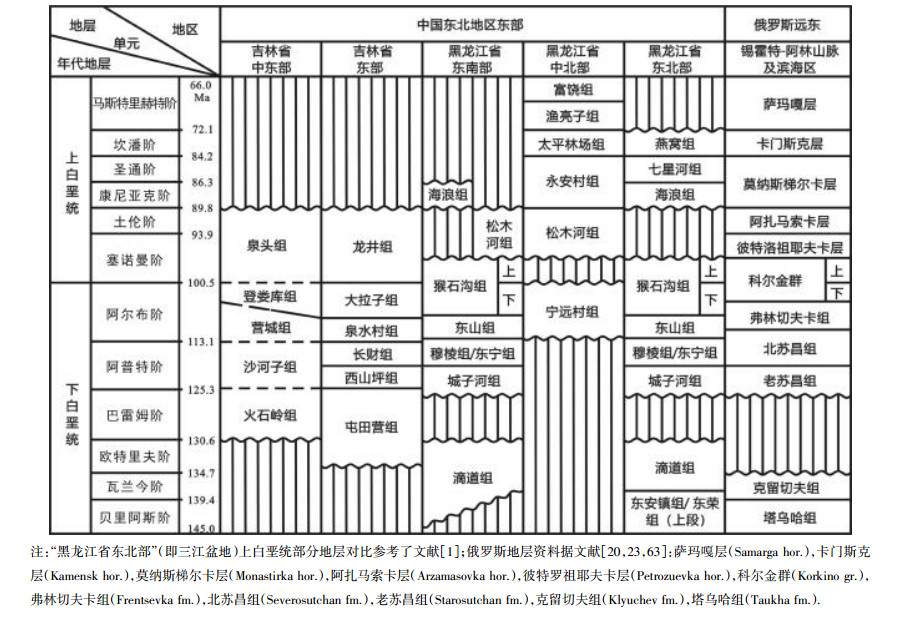
特别值得提及的是,前不久俄罗斯学者在俄南滨海苏昌盆地(Sutchan Basin,现称Partizask Basin)的波手—卡门镇(Bolshoy Kamen)附近(图 3a)的早白垩世地层中发现了保存完好的鸡西早期被子植物的代表分子:如优美亚洲叶(Asiatifolium elegans)和羽叶鸡西叶(Jixia pinnatipartita)等[23](图 2d-h). 俄罗斯学者认为上述被子植物的时代为早白垩世晚期,其层位可能属于弗林切夫卡组(Frenchevka Formation);但笔者判断该层位也可能为老苏昌组(Starosuchan Formation,与我国鸡西城子河组相当)(表 1). 当然,此问题还有待进一步研究和校正. 鸡西早期被子植物在俄罗斯南滨海地区的新发现,刷新了我们对鸡西早期被子植物古地理分布及发展演化的认识.

|
图 3 大拉子植物群及地层 Fig.3 Dalazi flora and strata a—俄罗斯苏昌盆地地理位置简图(据文献[24],本文加中文名并补充大拉子点位)(sketch map of Sutchan Basin in Russia,from Reference[23]);b—“中白垩纪”被子植物系统演化图(据文献[29])(evolution diagram of angiosperm in Middle Cretaceous,from Reference[29]);c—多国专家在大拉子组上段野外考察(1995)(international experts in field work on the U. mem. of Dalazi fm.);d,e—笔者课题组采集的大拉子组部分植物化石❶(some plant fossils of Dalazi flora collected by author’s research team);d—裸子植物及蕨类化石(gymnosperms and ferns):1—查米叶(Zamiophyllum),2,3—耳羽叶(Otozamites),4—小威廉姆逊(Williamsoniella),5—坚叶杉(Pagiophyllum),6—似里白(Gleichenites),7,8—枞型枝(Elatocladus);e—被子植物(angiosperms):9—拟蝙蝠葛(Menisphermites),10—延吉叶?(Yanjiphyllum?),11,12—苹婆叶(Sterculaephyllum),13,14—被子植物(未定名)(angiosperm,indet.) |
吉林东部延吉大拉子组早期被子植物的总体面貌较接近于以美国波托马克群(Potomac Group)早期被子植物为标准的演化序列第Ⅱ带(Zone-Ⅱ),时代为阿尔布期的中晚期(middle-late Albian)[29]❶(图 3b). 古植物学家陶君容和张川波最早较详细地研究了大拉子早期被子植物化石,认为其时代为阿普特—阿尔布期[30-31]. 自1986年起,笔者率课题组开始对大拉子组及其早期被子植物研究(图 3c-e),但对大拉子早期被子植物时代的认识,基本上沿用了陶君容和张川波的观点[10, 32].
❶孙革. 中生代植物. 沈阳师范大学研究生教材. 2019:1-81.
近年来,随着笔者团队与俄罗斯开展合作,有关大拉子组及其早期被子植物与俄罗斯南滨海地区具海陆交互相的白垩纪相关地层及化石的对比研究不断深入(图 3a). 俄罗斯南滨海的苏昌盆地早白垩世弗林切夫卡组(Frentsevka Formation)在植物群特征及地层层序等方面均可与大拉子植物群及其地层相比较. 但弗林切夫卡组具海相夹层,所产的海相双壳类三角蚌(Trigonia)指示中阿尔布期(middle Albian),因此弗林切夫卡组时代被认为属阿尔布期的中期或中晚期[20, 33-34]. 另一个证据是,弗林切夫卡组之上被科尔金群(Korkino Group)的下段整合覆盖,该下段的孢粉化石指示阿尔布晚期[20, 34]. 此外,弗林切夫卡组之下分别为含煤的北苏昌组(Severosutchan Formation,大体相当于我国的穆棱组)和含煤的老苏昌组(Starosutchan Formation,大体相当于我国的城子河组)(表 1). 据此,笔者综合多方面的资料认为:大拉子早期被子植物的时代可能为早白垩世阿尔布期的中晚期(middle-late Albian). 当然,有关大拉子组与俄罗斯南滨海弗林切夫卡组的详细对比还有待进一步工作.
值得提及的是,前不久,李想等[35]在对吉林大拉子组建组剖面研究中,获得大拉子组下段年龄为109.9±2.9 Ma,上段年龄为96.2±1.7 Ma,据此提出大拉子组的时代可能为早白垩世阿尔布期至晚白垩世塞诺曼期(Albian-Cenomanian). 笔者认为,上述大拉子组“上段”的地层及其测年结果还有待进一步讨论.
总之,大拉子早期被子植物是研究中国早期被子植物演化历程中的一个重要环节,也是与北美波托马克被子植物群对比的一个“窗口”. 有关大拉子早期被子植物的组成及时代研究还有待进一步深入.
2 晚白垩世植物群及地层研究进展 2.1 嘉荫晚白垩世植物新发现近年来,中国东北地区不断有晚白垩世植物化石发现,如在黑龙江嘉荫发现被子植物“黑龙江达勒比叶”(Dalembia heilongjiangensis)[36]及水生被子植物“嘉荫莲”(Nelumbo jiayinensis)[37]等,此外还发现水生被子植物“葛赫叶”(Quereuxia)的表皮构造[38]等(图 4b-i).

|
图 4 晚白垩世植物群研究进展 Fig.4 Advances in the study of Late Cretaceous flora a—嘉荫沿黑龙江晚白垩世地层出露示意图(outcrop of Upper Cretaceous strata in Jiayin along Heilongjiang River);b—黑龙江达勒比叶(Dalembia heilongjiangensis);c—达勒比叶在东北亚古地理分布图(据文献[36])(paleogeographic distribution of Dalembia in Northeast Asia,from Reference[36]);d,e—嘉荫莲(Nelumbo jiayinensis);f—嘉荫莲古生态复原图(据文献[37])(paleoecological restoration of Nelumbo jiayinensis,from Reference[37]);g,h,i—葛赫叶及其叶表皮细胞新发现(据文献[38])(new discovery of Quereuxia and its leaf epidermal cells,from Reference[38]);j,k,l—黑龙江悬铃木及化石产地(据文献[39, 41])(Platanus heilongjiangensis and fossil site,from Reference[39, 41]) |
嘉荫的达勒比叶(Dalembia)是东北亚地区白垩纪重要被子植物化石,在我国属首次发现. 由于在嘉荫发现的达勒比叶是该属在东北亚地区时代最晚、地理位置最南的代表,此发现扩大了对该属地质地理分布的认知(图 4 b、c). 嘉荫莲的发现曾引起轰动:因为“莲”即荷花,是我国“十大名花”之一,现今多生长在暖温带,此次莲化石在嘉荫的发现对恢复80 Ma前的嘉荫古气候及古环境提供了宝贵的化石依据;特别是此发现又将我国莲化石的地质记录提前了至少30 Ma [37](图 4d-f). 此外,水生被子植物化石角质层很难保存,其表皮细胞研究也较为罕见,梁飞等[38]首次发现葛赫叶的表皮细胞,是此领域研究的新突破(图 4g-i).
2.2 牡丹江植物组合及悬铃木类化石新发现牡丹江地区白垩纪被子植物化石最早由古植物学家张志诚首次发现[40]. 近年来,笔者等在牡丹江东北的七台河地区猴石沟组上段砂岩中又新发现悬铃类被子植物“黑龙江悬铃木”(Platanus heilongjiangensis)等化石;并首次获得了含化石层位整合上覆的火山岩96.2±1.7 Ma的同位素年龄,佐证了牡丹江植物组合的时代可能主要为晚白垩世塞诺曼期(Cenomanian). 这一新进展为建立中国东北地区白垩纪被子植物发展(演化)序列提供了新资料(图 4i-l)[39, 41].
牡丹江地区白垩纪被子植物化石,以往还发现有被子植物亚显赫悬铃木(Platanus subnobilis),假奎列尔玛悬铃木(P. pseudiguillelmae)及牡丹江惚木(Aralia mudanjiangensis)等[40]. 在其同期地层中还见有黑龙江崖柏(Thuja heilongjiangensis)等松柏类及少量蕨类等[42]. 此次黑龙江悬铃木的发现及同位素时代佐证,为牡丹江晚白垩世植物组合的确认提供了宝贵的证据. 由此,对黑龙江省东部晚白垩世植物群发展序列已大体识别出4个阶段:1)早期阶段——“牡丹江植物群(组合)”,以含大量被子植物悬铃木类为特色,时代约为Cenomanian期,气候总体上显示高热;2)“中期”阶段,主要为土伦—康尼亚克期(Turonian-Coniacian),其面貌目前尚不清楚,化石有待找寻;3)中晚期阶段——嘉荫植物群,杉科及银杏类繁盛,被子植物约占50%,总体上显示暖温带气候,时代为圣通—坎潘期(Santonian-Campnian);4)最晚期阶段——“渔—富植物群”(包括渔亮子组及富饶组组合),孢粉已显示富饶组早期组合中买麻藤类增加,晚期组合中被子植物已出现桦科等偏温凉植物,植物群已具一定的温带成分,显示气候已总体变凉,时代为马斯特里赫特期(Maastrichtian)[39].
2.3 晚白垩世地层新知 2.3.1 渔亮子组“二分”的发现近年来,嘉荫地区晚白垩世地层研究最重要的成果之一,是俄罗斯孢粉学家Markevich等[43]依据详细的孢粉研究将渔亮子组划分为上下两段(图 5a):下段以龙骨山层为代表,含满洲龙—卡龙动物组合,时代为Maastrichtian早期(图 5g-i);上段以乌拉嘎层为代表,含乌拉嘎龙—黑龙动物组合,时代为Maastrichtian中期(图 5b-f)[44-45].

|
图 5 黑龙江嘉荫晚白垩世地层及K—Pg界线研究进展 Fig.5 Advances in the study of Upper Cretaceous strata and K—Pg boundary in Jiayin, Heilongjiang Province a—嘉荫晚白垩世地层表(stratigraphic table of Upper Cretaceous in Jiayin);b~f—渔亮子组上段剖面(在乌拉嘎)及其化石(section of U. mem. of Yuliangzi fm. in Wulaga):c—乌拉嘎龙(Wulagasaurus),d—黑龙(Sahalyania),e—恐龙皮肤化石(dinosaur skin fossil),f—上段恐龙埋藏情况(burial of dinosaur in the U. mem.);g~i—渔亮子组下段剖面(在龙骨山)及其化石(section of L. mem. of Yuliangzi fm. in Longgushan):h—满洲龙(Mandschurosaurus),i—卡龙(Charonosaurus)(a~i据文献[45])(a-i from Reference[45]);j~l—嘉荫K—Pg界线的确定(据文献[44-45])(definition of K-Pg boundary,from References[44-45]);j—地层柱状示意图(stratigraphic column);k—确定界线的重要孢粉化石:下为“三棱鹰粉—条纹假鹰粉组合”(组合Ⅴ,产于富饶组),上为“混杂三孔庭粉—小刺鹰粉组合”(组合Ⅵ,产于乌云组白山头段)(significant sporopollen fossil for definition of boundary:the lower showing assemblage Ⅴ from Furao fm. and the upper assemblage Ⅵ from Baishantou mem. of Wuyun fm.);l— K-Pg界线之下的马斯特里赫特期的标准孢粉化石(Maasterichtian index fossils):1—三棱鹰粉(Aquilapollenites conatus),2—美丽等体粉(Intergricorpus bellum),3—清晰网假等体粉(Pseudointegricorpus clarireticulatus),4—斯氏鹰粉(Aquilapollenites stelkii),5—白垩小泡粉(Marsypiletes cretacea),6—粗糙沃氏粉(Wodehouseia aspera) |
嘉荫地区及其北部邻区晚白垩世恐龙已查明至少由9个分类群组成,但以往长期笼统认为这里的恐龙的时代是马斯特里赫特期[46-48],缺少详细的时代划分,这便在一定程度上影响了对这里的恐龙发展及灭绝时间的认识. 近年来,经Markevich等[43]详细研究,确认渔亮子组下段(含龙骨山恐龙动物组合)产孢粉化石“阿米格黛鹰粉-明确买麻藤粉组合”(Aquilapollenites amygdaloides-Gnetaceaepollenites evidens Ass.,组合Ⅲ),时代为马斯特里赫特早期;渔亮子组上段(含乌拉嘎恐龙动物组合)孢粉化石为“粗糙沃氏粉—黑龙江小突粉组合”(Wodehouseia aspera-Parviprojectus amurensis Ass.,组合Ⅳ),时代为马斯特里赫特中期. 这一详细的时代划分表明,嘉荫恐龙鼎盛时期是以龙骨山的满洲龙、卡龙为代表的马斯特里赫特早期;到了乌拉嘎时期(以乌拉嘎龙、黑龙等为代表),时代为马斯特里赫特中期,嘉荫恐龙已走向衰落[44-45];到富饶组(马斯特里赫特晚期)已不见恐龙踪迹,似说明嘉荫恐龙可能在马斯特里赫特中期之末已经灭绝(图 5j). 也就是说,早在K—Pg界线之前,嘉荫恐龙已经绝灭[45]. 此外,笔者团队研究表明:嘉荫地区K—Pg界线附近地层并没有发现“铱异常”,表明此间在嘉荫地区并没有“小行星撞击”事件发生;嘉荫乃至整个东北亚地区,晚白垩世晚期的火山活动、海平面下降和气候变冷等可能是嘉荫恐龙动物群灭绝的主要原因[44-45, 49-50].
2.3.2 黑龙江嘉荫陆相K—Pg界线的确立白垩纪—古近纪地层(K—Pg)界线,是发生在距今66 Ma的一条地层界线. 该界线出现前后,地球生物与环境曾发生重大变化:包括恐龙在内的70%以上的陆生生物灭绝、海洋中以菊石为代表的大多数生物也同时消亡. 这一事件被称为“显生宙以来的第5次生物大灭绝与复苏事件”. 由此,有关K—Pg界线研究也成为国际前沿课题之一[51-52].
我国以往在此领域研究较薄弱. 自2002年起,在国家自然科学基金重大国际项目等支持下,笔者率领由中、俄、德、美、英、比、日等多国科学家组成的国际科研队,通过长达10年的潜心研究,运用古生物学、地球化学及古地磁学等综合研究方法,并实施3个钻孔,以孢粉研究为“龙头”,最终于2011年在嘉荫小河沿首次确定了中国陆相K—Pg界线[44-45, 53-59]. 2011年8月,来自中、俄、德、美、英、法、日、韩、印度等15国近百名专家出席了在嘉荫举行的K—Pg界线立碑典礼,一致认定嘉荫小河沿的K—Pg界线是“中国首个具有可靠证据的陆相白垩纪—古近纪地层界线”. 2019年8月来自16国百余名专家又齐聚嘉荫,再次确认并建议嘉荫小河沿的K—Pg界线作为“中国陆相K—Pg界线的国家标准”,全国地层委员会也为此发来贺信. 目前,该界线点已被确定为国际陆相K—Pg界线第95号候选点[52].
嘉荫小河沿的K—Pg界线位于XHY—2006钻孔的22.00~22.05 m的地层之间:界线之下产有典型晚白垩世马斯特里赫特晚期“三棱鹰粉—清晰网假等体粉(Aquilapollenites stelkii-Pseudointegricorpus clarireticulatus)”孢粉组合(组合Ⅴ);界线之上有典型古新世丹表早期的“混杂三孔庭粉—小刺鹰粉(Triatriopollenites confusus-Aquilapollenites spinulosus)”孢粉组合(组合Ⅵ)[43-45, 58](图 5k、l). K—Pg界线之上还有酸性火山岩64.1±0.7 Ma等同位素测年证据[59],界线之下有66.3±1.7 Ma的测年证据等❶;古地磁测试结果也支持此界线划定的准确性[44]. 所有这些为K—Pg界线的确定提供了可靠证据.
❶孙革,等. 中国白垩系-古近系界线研究(“中国标准地层建立——中国地层表完善”最终报告).科技部科技基础专项项目(2015FY-310100). 2022:1-65.
近年来,笔者率课题组承担科技部专项项目“中国白垩系—古近系界线研究”(2015—2018),对嘉荫沿江白垩纪地层剖面开展了新一轮调查研究工作;冯玉辉等已在永安村组下伏的火山岩测得99.5 Ma的年龄❶.后者可能相当于松木河组(含Cenomanian期). 由此,黑龙江嘉荫的沿江白垩系剖面有望成为一条出露较完整的上白垩统剖面.
3 嘉荫晚白垩世动、植物的协同演化 3.1 恐龙的“餐桌”白垩纪植物是这一时期植食性恐龙的主要食物来源,而植食性恐龙又是肉食性恐龙食物的一部分. 因此,植物界的繁荣与否关系到恐龙的繁衍生息,而恐龙的繁盛与否也是该时期植物界发展状况的“晴雨表”. 植食性恐龙包括所有鸟臀类和大部分蜥臀类恐龙,它们通常身体庞大:如中国河南晚白垩世的汝阳龙(Ruyangsaurus)体长已达38 m,如此庞大的身体通常需要有足够的食物供给. 从形态功能特征看,嘉荫晚白垩世植食性恐龙通常形体较高,大都颈部较长,易于向高处取食,如满洲龙(Mandschurosaurus)、卡龙(Charonosaurus)等身长达11 m,颈部长至少4~5 m;大天鹅龙(Olorotitan)身长约8 m,颈部由18根颈骨组成. 所有这些反映它们的身体与被取食的植物的高度(如杉科植物通常都高达30 m以上)相匹配. 从恐龙的牙齿看:植食性恐龙的牙齿多为勺形齿或钉状齿,便于剪断茎叶或刺穿球果外壳,似说明当时的植被可能已具球果等(如松柏类);特别是,嘉荫地区鸭嘴龙类恐龙的上、下颌牙齿呈多行排列(如大天鹅龙具齿32列),牙齿数百颗,这些牙齿便于对果实的研磨(图 6a、b);蕨类植物也应是恐龙的主要食物之一,其过多的消耗可能是自中生代中晚期起蕨类出现逐渐“矮化”的原因之一[39](图 6c-e).

|
图 6 嘉荫晚白垩世植物群与恐龙协同演化及“恐龙餐桌” Fig.6 The co-evolution of Late Cretaceous flora and dinosaurs in Jiayin and plants serving as food for dinosaur a,b—鸭嘴龙类下颌及牙齿(据文献[45])(lower jaw and teeth of hadrosaurid,from Reference[45]);c,d,e—嘉荫及其邻区晚白垩世鸭嘴龙类及生态复原图(Late Cretaceous hadrosaurid and ecological restoration in Jiayin and neighboring area):d—阿穆尔龙(Amurosaurus),e—昆都尔龙(Kundurosaurus);f—鹦鹉嘴龙食苏铁类复原(restoration of Psittacosaurus eating cycads);g—鸭嘴龙类与水生被子植物古生态复原(据文献[60])(paleoecological restoration of hadrosaur with aquatic angiosperms,from Reference[60]);h—水生被子植物葛赫叶(嘉荫,K2)(aquatic angiosperm Quereuxia,Jiayin);i~o—现生植物(living plants):i—蕨类(fern),j,k—苏铁(Cycas),l—银杏(Ginkgo),m—红豆杉(Taxus),n—松球果(conse of Pinus),o—木兰(Magnolia) |
那么,究竟哪些植物曾上了植食恐龙的“餐桌”呢?显然,蕨类植物应是首选,它们分布广,易于获取,叶子细嫩,易于恐龙消化;银杏类的果子和苏铁类的大孢子叶(如苏铁果等)也是香甜可口的食物;松柏类杉科(如水杉、红杉等)叶子和球果也十分丰富;大量被子植物,特别是水生被子植物(如葛赫叶、卡波叶及莲等)叶子柔软,又暴露在水面,会受恐龙青睐(图 6f-l,n,o). 前不久,有报道在内蒙古固阳早白垩世地层中已发现“红豆杉”(Taxus)化石[61],红豆杉“果子”甜美(图 6m),到晚白垩世这类植物会更受恐龙喜欢[39].
从近年来新发现看,嘉荫晚白垩世中晚期植物已至少发现34属43种大化石,其中蕨类约占12%,银杏类约7%,松柏类约28%,被子植物约占50% [39]. 繁茂的杉科植物(如水杉Metasequoia、红杉Sequoia)、银杏类、大量喜湿的蕨类及丰富的被子植物等,都可能会为恐龙的“餐桌”提供了保障.
3.2 晚白垩世中—晚期恐龙与嘉荫植物的会聚在中国的恐龙分布简图上,可以看到恐龙“大家庭”的一个“主支”在晚白垩世最终迁徙到我国东北地区黑龙江嘉荫一带,并在这里最终消亡(图 7a). 恐龙的迁移通常与植物群发展(迁移)方向一致,这也是动、植协同演化在宏观上的反映. 恐龙主要生活在森林地区或植物繁茂的开阔地带. 晚白垩世中晚期的嘉荫地区(包括俄罗斯结雅—布列亚盆地)气候温暖湿润、植被繁茂,曾吸引了众多的恐龙迁徙到这里. 经近年来研究,嘉荫的植食性恐龙占95%以上,已发现至少9个分类群[45-48],鸭嘴龙类包括满洲龙、卡龙、克伯龙(Kerberosaurus)、乌拉嘎龙(Wulagasaurus)、黑龙(Sahaliyania)、昆都尔龙(Kundurasaurus)、阿穆尔龙(Amurosaurus)及大天鹅龙,还有嘉荫龙足印(Jiayinosauropis)反映的恐龙[54]等. 当然,以暴龙类为代表的肉食性恐龙也在此繁衍[60](图 7b-d). 自晚白垩世中期的圣通期至晚白垩世晚期的马斯特里赫特早中期,由于植物界的繁茂,嘉荫的恐龙曾一度繁盛;但到马斯特里赫特晚期,由于火山活动等影响,加之嘉荫地区气候可能变凉,该地区的植被已走向衰退,可能已无力再支持恐龙的繁衍和生存. 至此,植食性恐龙可能首先消亡,之后,肉食性恐龙也随之殆尽. 这一“悲剧”为思考动植物的协同演化、乃至恐龙灭绝的故事提供了参考[50].

| [1] |
黑龙江省地质矿产局. 黑龙江省区域地质志[M]. 北京: 地质出版社, 1993: 1-734. Heilongjiang Bureau of Geology and Mineral Resources. Regional geology of Heilongjiang Province[M]. Beijing: Geological Publication House, 1993: 1-734. |
| [2] |
沙金庚, 袁福盛. 黑龙江饶河东安镇组的Buchia化石群[J]. 古生物学报, 1985, 24(6): 651-662. Sha J G, Yuan F S. Buchia fauna from Dong'anzhen Formation near Dong'an of Raohe County, Heilongjiang[J]. Acta Palaeontologica Sinica, 1985, 24(6): 651-662. |
| [3] |
孙革, 沙金庚, 王义刚, 等. 黑龙江饶河东安镇侏罗-白垩系界线及Buchia新知[J]. 古生物学报, 1989, 28(5): 579-590, 597. Sun G, Sha J G, Wang Y G, et al. Jurassic-Cretaceous boundary of Dong'anzhen, Raohe, Heilongjiang with a new knowledge of Buchia[J]. Acta Palaeontologica Sinica, 1989, 28(5): 579-590, 597. |
| [4] |
孙革, 郑少林, 孙学坤, 等. 黑龙江东部侏罗-白垩系界线附近地层研究新进展[J]. 地层学杂志, 1992, 16(1): 49-54. Sun G, Zheng S L, Sun X K, et al. Recent advance in the research of the strata near the Jurassic-Cretaceous boundary in eastern Heilongjiang[J]. Journal of Stratigraphy, 1992, 16(1): 49-54. |
| [5] |
余静娴. 黑龙江省绥滨地区晚侏罗世晚期-早白垩世沟鞭藻组合[C]//中国地质科学院沈阳地质矿产研究所所刊. 沈阳: 辽宁科学技术出版社, 1982, 5: 227-267. Yu J X. Late Jurassic and Early Cretaceous dinoflagellate assemblages of eastern Heilongjiang Province, China[C]//Bulletin of the Shenyang Institute of Geology and Mineral Resources, Chinese Academy of Geological Sciences. Shenyang: Liaoning Science and Technology Publishing Houses, 1982, 5: 227-267. |
| [6] |
席党鹏, 万晓樵, 李国彪, 等. 中国白垩纪综合地层和时间框架[J]. 中国科学: 地球科学, 2019, 62(1): 257-288. Xi D P, Wan X Q, Li G B, et al. Cretaceous integrative stratigraphy and timescale of China[J]. Science China Earth Sciences, 2019, 62(1): 256-286. |
| [7] |
程金辉, 何承全. 黑龙江省东部鸡西盆地早白垩世滴道组的沟鞭藻类[J]. 古生物学报, 2001, 40(1): 127-133. Cheng J H, He C Q. Early Cretaceous dinoflagellates from the Didao Formation in the Jixi Basin, eastern Heilongjiang Province, NE China[J]. Acta Palaeontologica Sinica, 2001, 40(1): 127-133. DOI:10.3969/j.issn.0001-6616.2001.01.012 |
| [8] |
Zhu Y H, He C Q. Middle Jurassic to Early Cretaceous dinoflagellate assemblage zones in eastern Heilongjiang Province, Northeast China[J]. Cretaceous Research, 2007, 28(2): 327-332. DOI:10.1016/j.cretres.2006.10.001 |
| [9] |
郑少林, 张武. 黑龙江省东部地区龙爪沟群及鸡西群植物化石[C]//中国地质科学院沈阳地质矿产研究所所刊. 沈阳: 辽宁科学技术出版社, 1982, 5: 277-382. Zheng S L, Zhang W. Plant fossils of Longzhaogou and Jixi groups in eastern Heilongjiang Province[C]//Bulletin of the Shenyang Institute of Geology and Mineral Resources, Chinese Academy of Geological Sciences. Shenyang: Liaoning Science and Technology Publishing House, 1982, 5: 277-382. |
| [10] |
孙革, 曹正尧, 李浩敏, 等. 白垩纪植物群[C]//李星学. 中国地质时期植物群. 广州: 广东科技出版社, 1995: 310-341. Sun G, Cao Z Y, Li H M, et al. Cretaceous floras of China[C]//Li X X. Fossil floras of china through the geological ages. Guangzhou: Guangdong Science & Technology Press, 1995: 310-341. (in Chinese) |
| [11] |
Geyler H T. Ueber fossile Pflanzen aus der Juraformation Japans[J]. Palaeontographica, 1877, 24(5): 221-232. |
| [12] |
Kimura T. The present status of the Mesozoic land flora of Japan[M]. Tsukuba: Tsukuba University Press, 1980: 379-413.
|
| [13] |
Kusuhashi N, Matsumoto A, Murakami M, et al. Zircon U-Pb ages from tuff beds of the Upper Mesozoic Tetori Group in the Shokawa district, Gifu Prefecture, central Japan[J]. Island Arc, 2006, 15(3): 378-390. DOI:10.1111/j.1440-1738.2006.00544.x |
| [14] |
Legrand J, Yamada T, Terada K, et al. Palynofloras from the Itsuki and Kuwajima formations of the Tetori Group and their correlation with Paleofloristic Provinces of Eastern Asian[J]. Paleontological Research, 2021, 25(3): 177-190. |
| [15] |
Feng Y H, Yang T, Liang F, et al. New zircon U-Pb age of the Didao Formation in Jixi Basin and its significance for the geology and paleogeography in Jixi and eastern Heilongjiang region in the Early Cretaceous[J]. Cretaceous Research, 2022, 135: 105169. DOI:10.1016/j.cretres.2022.105169 |
| [16] |
孙革, 郭双兴, 郑少林, 等. 世界最早的被子植物化石群的首次发现[J]. 中国科学B辑, 1992, 22(5): 543-548. Sun G, Guo S X, Zheng S L, et al. Discovery of the earliest known angiosperms in the world[J]. Science in China (Series B), 1992, 22(5): 543-548. |
| [17] |
Sun G, Dilcher D L. Early angiosperms from Lower Cretaceous of Jixi, China and their significance for study of the earliest occurrence of angiosperms in the world[J]. Palaeobotanist, 1996, 45: 393-399. |
| [18] |
Sun G, Dilcher D L. Early angiosperms from the Lower Cretaceous of Jixi, eastern Heilongjiang, China[J]. Review of Palaeobotany and Palynology, 2002, 121(2): 91-112. DOI:10.1016/S0034-6667(02)00083-0 |
| [19] |
孙革, 刘风香. 黑龙江鸡西含煤盆地早白垩世城子河组特尔姆叶(Tyrmia)的发现[J]. 煤田地质与勘探, 2007, 35(2): 1-5. Sun G, Liu F X. Discovery of Tyrmia from Lower Cretaceous Chengzihe Formation in Jixi coal-bearing basin of Heilongjiang, China[J]. Coal Geology & Exploration, 2007, 35(2): 1-5. DOI:10.3969/j.issn.1001-1986.2007.02.001 |
| [20] |
Volynets E. Geology and forming condition of Aptian-Cenimanian deposits in NW Primorye[D]. IBSS FEB RAS, 2011.
|
| [21] |
孙革, 郑少林, 姜剑红, 等. 黑龙江鸡西含煤盆地早白垩世生物地层研究新进展[J]. 煤田地质与勘探, 1999, 27(6): 1-3. Sun G, Zheng S L, Jiang J H, et al. Recent advance in study of Early Cretaceous biostratigraphy from Jixi coal-bearing basin, Heilongjiang, China[J]. Coal Geology & Exploration, 1999, 27(6): 1-3. DOI:10.3969/j.issn.1001-1986.1999.06.001 |
| [22] |
孙革, 郑少林. 中国东北中生代地层划分对比之新见[J]. 地层学杂志, 2000, 24(1): 60-64. Sun G, Zheng S L. New proposal on division and correlation of Mesozoic from northeastern China[J]. Journal of Stratigraphy, 2000, 24(1): 60-64. DOI:10.3969/j.issn.0253-4959.2000.01.009 |
| [23] |
Golovneva L, Alekseev P, Bugdaeva E, et al. An angiosperm dominated herbaceous community from the Early-Middle Albian of Primorye, Far East of Russia[J]. Fossil Imprint, 2018, 74(1/2): 165-178. |
| [24] |
Krassilov V, Volynets E. Weedy Albian angiosperms[J]. Acta Palaeobotanica, 2008, 48(2): 151-169. |
| [25] |
沙金庚. 黑龙江省东部Aucellina (双壳类)的发现[J]. 地层学杂志, 1990, 14(3): 226-230. Sha J G. Discovery of Aucellina (bivalves) in eastern Heilongjiang[J]. Journal of Stratigraphy, 1990, 14(3): 226-230. |
| [26] |
沙金庚. 黑龙江东部早白垩世生物地层学研究的主要进展[J]. 地学前缘, 2002, 9(3): 95-101. Sha J G. Major achievements in studying the Early Cretaceous biostratigraphy of eastern Heilongjiang[J]. Earth Science Frontiers, 2002, 9(3): 95-101. DOI:10.3321/j.issn:1005-2321.2002.03.013 |
| [27] |
Sha J G. Cretaceous stratigraphy of northeast China: Non-marine and marine correlation[J]. Cretaceous Research, 2007, 28(2): 146-170. DOI:10.1016/j.cretres.2006.12.002 |
| [28] |
何承全, 孙学坤. 黑龙江省东部鸡西盆地城子河组下部早白垩世欧特里夫晚期海相沟鞭藻类[J]. 古生物学报, 2000, 39(1): 46-62. He C Q, Sun X K. Late Hauterivian dinoflagellates from the lower part of the Chengzihe Formation in Jixi Basin, eastern Heilongjiang, NE China[J]. Acta Palaeontologica Sinica, 2000, 39(1): 46-62. DOI:10.3969/j.issn.0001-6616.2000.01.003 |
| [29] |
Doyle J A. Molecular and fossil evidence on the origin of angiosperms[J]. Annual Review of Earth and Planetary Sciences, 2012, 40: 301-326. DOI:10.1146/annurev-earth-042711-105313 |
| [30] |
陶君容, 张川波. 吉林省延吉盆地早白垩世被子植物化石[J]. 植物学报, 1990, 32(3): 220-229. Tao J R, Zhang C B. Early Cretaceous angiosperms of the Yanji Basin, Jilin Province[J]. Acta Botanica Sinica, 1990, 32(3): 220-229. |
| [31] |
陶君容, 张川波. 中国早白垩纪被子植物生殖器官[J]. 植物分类学报, 1992, 30(5): 423-426. Tao J R, Zhang C B. Two angiosperm reproductive organs from the Early Cretaceous of China[J]. Acta Phytotaxonomica Sinica, 1992, 30(5): 423-426. |
| [32] |
Sun G, Zheng S L, Wang X F, et al. Subdivisions of main developmental stages of early angiosperms in NE China[J]. Acta Palaeontologica Sinica, 2000, 39(S): 186-199. |
| [33] |
Krassilov V A. Early Cretaceous flora of Southern Primorye and its significance for stratigraphy[M]. Moscow: Nauka, 1967: 1-167.
|
| [34] |
Krassilov V A, Nevolina S I, Philinnova G G. Development of floras from the Far East and geological events in the Middle Cretaceous[C]//IGCP 58, IBSS, FEB AS USSR. Vladivostok, 1981: 103-115.
|
| [35] |
李想, 高妍, 张德军, 等. 吉林延边智新盆地大拉子组地质时代: 来自锆石U-Pb年代学证据[C]//中国古生物学会第28届学术年会论文摘要集. 沈阳: 中国古生物学会, 2015: 165-166. Li X, Gao Y, Zhang D J, et al. The geological age of Dalazi Formation from Zhixing Basin of Yanbian, Jilin: Evidenced by U-Pb zircons dating[C]//Abstracts of the 28th Annual Symposium of Paleotological Society of China. Shenyang: Palaeontological Society of China, 2015: 165-166. (in Chinese) |
| [36] |
Sun G, Golovneva L, Alekseev P, et al. New species Dalembia jiayinensis (Magnoliopsida) from the Upper Cretaceous Yong'ancun Formation, Heilongjiang, northern China[J]. Cretaceous Research, 2016, 67: 8-15. DOI:10.1016/j.cretres.2016.06.011 |
| [37] |
Liang F, Sun G, Yang T, et al. Nelumbo jiayinensis sp. nov. from the Upper Cretaceous Yong'ancun Formation in Jiayin, Heilongjiang, Northeast China[J]. Cretaceous Research, 2018, 84: 134-140. DOI:10.1016/j.cretres.2017.11.007 |
| [38] |
Liang F, Tian N, Sun W, et al. Epidermal features of the floating leaves of Quereuxia angulata (Newberry) Krištofovič, an aquatic angiosperm from the Upper Cretaceous of Northeast China[J]. Cretaceous Research, 2021, 125: 104835. DOI:10.1016/j.cretres.2021.104835 |
| [39] |
孙革, 梁飞, 杨涛, 等. 黑龙江嘉荫晚白垩世植物群[M]. 上海: 上海科技教育出版社, 2020: 1-176. Sun G, Liang F, Yang T, et al. Late cretaceous flora from Jiayin of Heilongjiang, China[M]. Shanghai: Shanghai Science Technology Education Press, 2020: 1-176. |
| [40] |
张志诚. 牡丹江盆地的几种白垩纪被子植物化石[C]//中国地质科学院院报: 沈阳地质矿产研究所分刊. 北京: 地质出版社, 1981, 2(1): 154-162. Zhang Z C. Several Cretaceous angiospermous from Mudanjiang Basin, Heilongjiang[J]. Bulletin of the Chinese Academic Geological Science. Beijing: Geological Publishing House, 1981, 2(1): 154-162. |
| [41] |
Sun G, Kovaleva T, Liang F, et al. A new species of Platanus from the Cenomanian (Upper Cretaceous) in eastern Heilongjiang, China[J]. Geoscience Frontiers, 2019, 10(4): 1535-1541. DOI:10.1016/j.gsf.2018.10.006 |
| [42] |
郑少林, 张莹. 松辽盆地的白垩纪植物[J]. 古生物学报, 1994, 33(6): 756-764. Zheng S L, Zhang Y. Cretaceous plants from Songliao Basin, Northeast China[J]. Acta Palaeontologica Sinica, 1994, 33(6): 756-764. |
| [43] |
Markevich V S, Sun G, Ashraf A R, et al. The Maastrichtian-Danian palynological assemblages from Wuyun of Jiayin nearby the Heilongjiang (Amur) River[C]//Yang Q, Yang Y D, Weldon E A. Ancient life and modern approaches. Hefei: University of Science and Technology of China Press, 2006: 526-527.
|
| [44] |
Sun G, Akhmetiev M, Markevich V, et al. Late Cretaceous biota and the Cretaceous-Paleogene (K-Pg) boundary in Jiayin of Heilongjiang, China[J]. Global Geology, 2011, 14(3): 115-143. |
| [45] |
孙革, 董枝明, 阿克米梯耶夫, 等. 黑龙江嘉荫晚白垩世-古新世生物群、K-Pg界线及恐龙灭绝[M]. 上海: 上海科技教育出版社, 2014: 1-196. Sun G, Dong Z M, Akhmetiev M, et al. Late Cretaceous-Paleocene biota and the K-Pg boundary from Jiayin of Heilongjiang, China with discussion on the extinction of dinosaurs[M]. Shanghai: Shanghai Science Technology Education Press, 2014: 1-196. |
| [46] |
Bolotsky Y L, Godefroit P. A new hadrosaurine dinosaur from the Late Cretaceous of Far Eastern Russia[J]. Journal of Vertebrate Paleontology, 2004, 24(2): 351-365. DOI:10.1671/1110 |
| [47] |
Godefroit P, Hai S L, Yu T X, et al. New hadrosaurid dinosaurs from the uppermost Cretaceous of northeastern China[J]. Acta Palaeontologica Polonica, 2008, 53(1): 47-74. DOI:10.4202/app.2008.0103 |
| [48] |
Godefroit P, Lauters P, Van Itterbeeck J, et al. Recent advances on study of hadrosaurid dinosaurs in Heilongjiang (Amur) River area between China and Russia[J]. Global Geology, 2011, 14(3): 160-191. |
| [49] |
Haq B U, Hardenbol J, Vail P R. Chronology of fluctuating sea levels since the Triassic[J]. Science, 1987, 235(4793): 1156-1167. DOI:10.1126/science.235.4793.1156 |
| [50] |
孙革, 董枝明, 吴文昊, 等. 中国最晚恐龙家族的消亡及黑龙江白垩纪-古近纪地层界线的确定[J]. 大自然, 2015(4): 28-31. Sun G, Dong Z M, Wu W H, et al. The extinction of the latest dinosaur fauna and the K-Pg boundary from Jiayin of Heilongjiang, China[J]. China Nature, 2015(4): 28-31. DOI:10.3969/j.issn.0255-7800.2015.04.006 |
| [51] |
Alvarez L W, Alvarez W, Asaro F, et al. Extraterrestrial cause for the Cretaceous-Tertiary extinction[J]. Science, 1980, 208(4448): 1095-1108. DOI:10.1126/science.208.4448.1095 |
| [52] |
Nichols D J, Johnson K R. Plants and the K-T boundary[M]. Cambridge: Cambridge University Press, 2008: 1-292.
|
| [53] |
Sun G, Akhmetiev M A, Dong Z M, et al. In search of the Cretaceous-Tertiary boundary in Heilongjiang River area of China[J]. Journal of Geoscience Research NE Asia, 2002, 5(2): 105-113. |
| [54] |
董枝明, 周忠立, 伍少远. 记黑龙江畔一鸭嘴龙足印化石[J]. 古脊椎动物学报, 2003, 41(4): 324-326. Dong Z M, Zhou Z L, Wu S Y. Note on a hadrosaur footprint from Heilongjiang River area of China[J]. Vertebrata Palasiatica, 2003, 41(4): 324-326. DOI:10.3969/j.issn.1000-3118.2003.04.007 |
| [55] |
Sun G, Akhmetiev M A, Golovneva L, et al. Late Cretaceous plants from Jiayin along Heilongjiang River, Northeast China[J]. Courier Forschungsinstitut Senckenberg, 2007, 258: 75-83. |
| [56] |
Quan C, Sun G. Late Cretaceous aquatic angiosperms from Jiayin, Heilongjiang, Northeast China[J]. Acta Geologica Sinica, 2008, 82(6): 1133-1140. |
| [57] |
Golovneva L B, Sun G, Bugdaeva E V. Campanian flora of the Bureya River Basin (Late Cretaceous of the Amur Region)[J]. Paleontological Journal, 2008, 42(5): 554-567. DOI:10.1134/S0031030108050110 |
| [58] |
Markevich V S, Bugdaeva E V, Ashraf A R, et al. Boundary of Cretaceous and Paleogene continental deposits in Zeya-Bureya Basin, Amur (Heilongjiang) River region[J]. Global Geology, 2011, 14(3): 144-159. |
| [59] |
Suzuki S, Sun G, Knittel U, et al. Radiometric zircon ages of a tuff sample from the Baishantou Member of Wuyun Formation, Jiayin: A contribution to the search for the K-T Boundary in Heilongjian River area, China[J]. Acta Geologica Sinica (English Edition), 2011, 85(6): 1351-1358. DOI:10.1111/j.1755-6724.2011.00591.x |
| [60] |
Stockey R A, Rothwell G W, Johnson K R. Cobbania corrugata gen. et comb. nov.(Araceae): A floating aquatic monocot from the Upper Cretaceous of western North America[J]. American Journal of Botany, 2007, 94(4): 609-624. DOI:10.3732/ajb.94.4.609 |
| [61] |
徐小慧, 杨柳荫, 孙柏年, 等. 早白垩世红豆杉属化石表皮构造的探索性研究[J]. 安徽理工大学学报(自然科学版), 2017, 37(2): 1-5. Xu X H, Yang L Y, Sun B N, et al. Exploratory study of the epidermal structure of the Early Cretaceous Taxus fossils[J]. Journal of Anhui University of Science and Technology (Natural Science), 2017, 37(2): 1-5. |
| [62] |
伊万·包罗斯基. 黑龙江(阿穆尔)地区晚白垩世暴龙类恐龙(虚骨龙类)[D]. 长春: 吉林大学, 2013. Bolotsky I. Tyrannosaurid dinosaurs (Coelurosauria) from Upper Cretaceous of Amur/Heilongjiang area[D]. Changchun: Jilin University, 2013. |
| [63] |
Mikhailov V A, Reibalko V I. Stratigraphy of eastern Shikhot-Alin volcanical region[C]//Volcanogenic Cretaceous of the Far East. IUC-UNESCO Project 245. Vladivostok. 1989: 14-35.
|