 2021, Vol.41
2021, Vol.41



2 青岛海洋科学与技术试点国家实验室海洋地质过程与环境功能实验室, 山东 青岛 266237;
3 中国科学院大学, 北京 100049;
4 中海辉固地学服务(深圳)有限公司, 广东 深圳 518068)
黄海是我国东部一个典型的半封闭陆架边缘海,位于中国大陆和朝鲜半岛之间,有着平坦宽阔、构造稳定的海底,从胶东半岛成山角到朝鲜长山串之间海面最窄(193km),习惯上以此连线将黄海分为北黄海和南黄海两部分[1]。作为中国东部边缘海的主要组成部分,黄海是黄河和长江等东亚大河的主要沉积汇[2]。虽然现今黄河与长江并不在黄海入海,但这两大河流仍很大程度上影响着并且在地质历史时期曾经直接控制着黄海的沉积环境及演化[3~6]。新近纪以来,南黄海南部和北部两个盆地的沉积物厚达1600m,中部隆起沉积较薄,约200~400m[1]。这些堆积于黄海盆地的巨厚陆源沉积物反过来也成为了重建地质历史时期高分辨率大陆古气候和古环境演变的极佳载体[6~9]。
河流沉积物中,除了常见的碎屑矿物和有机质外,还有一种含量极低而易被忽略的含碳物质,即黑碳(black carbon,通常 < 0.5 %)[10]。黑碳是生物质或化石燃料不完全燃烧形成的具有高度热稳定性的连续体,主要包括烟炱、木炭、焦炭和石墨碳[11]。黑碳大部分会储存在土壤中,一部分会随着风、地表径流搬运,最终在湖泊、海洋等环境中沉积下来[12]。对边缘海沉积物黑碳来源的研究表明,河流是黑碳由陆地进入海洋的主要途径[10, 13]。黑碳由于其稳定的化学结构,具有很高的抗氧化性和抗分解能力,加上其在沉积期后所受到的光化学作用和微生物作用很小,黑碳在沉积环境中可以保存百万年以上[14]。因此,黑碳的沉积记录可能包含了源区植被信息,是重建过去古环境、古气候演变的重要载体[15~16]。
自然界中陆生植物根据其光合作用途径的不同,分为C3型植物、C4型植物和景天酸代谢型植物(或称CAM型)三大类,其中所有的乔木、绝大多数灌木和草本植物使用C3光合作用途径(约85 %),而部分灌木和草本植物使用C4光合作用途径(约15 %),CAM型植物占比很少,主要为沙漠生存的耐干旱肉质植物(如仙人掌)[17]。C4光合作用途径的出现是新生代地球植物演化史上极其重要事件之一[18]。C4相对C3植物光合作用途径的优势在于“CO2浓缩机制”,其演化出的特殊叶片结构在羧化固定前可浓缩提高CO2的浓度,从而减弱了光呼吸作用。这种机制使得C4植物能够在低CO2浓度、高温、强光照、干旱和盐碱化等环境条件下仍具有较高的光合作用效率;而在同样的环境条件下,C3植物光呼吸作用强烈,光合作用效率明显降低[18]。C3、C4植物由于光合作用途径不同,碳的同位素分馏形成了差异,导致两者具有明显不同的δ13C值范围。C3植物δ13C的最高频率值为-26.7±2.3 ‰,而C4植物δ13C的最高频率值为-12.5±1.1 ‰ [19]。研究表明现代C4植物主要分布在温度相对较高的中低纬度区域和海拔相对较低的区域[20~21]。因此,C4植被的分布变化在一定程度上反映了古气候的演化[22]。而植被不完全燃烧产生的黑碳,其δ13C值很大程度上保留了原始植被的这种特征,这对重建黑碳源区的古植被、古气候演化历史具有重要意义[10]。
C4植被在地质历史时期曾发生过多次扩张,尤其在晚中新世被广泛认为发生了C4植被扩张[19~20, 22~30]。根据C4植物的生物学特性,其扩张可能受到了大气CO2浓度、干旱度、夏季风演化、降水季节性变化以及火活动的影响[19, 23, 27, 31~34]。目前中国C4植被演化研究多集中于黄土高原地区。灵台剖面土壤碳酸盐的碳同位素记录表明C4植被从约4Ma开始扩张,与低纬南亚地区相比滞后了约3Ma,而3Ma以来又明显减少,这不排除大气CO2浓度的降低可能引起了7~4Ma全球C4植被的扩张[35],但是高低纬度地区C4植被扩张的不同步性,表明除了CO2和温度因素以外还有其他区域或局部因素,如暖季降水,对C3/C4植被演化也有重要作用[36]。蓝田和灵台剖面的土壤碳酸盐和有机质碳同位素记录则表明,与季风演化相关的暖季降水和温度变化是半干旱地区7Ma以来C3/C4植被演化的主要原因,3.0~1.5Ma时期C4植被相对丰度的减少与夏季风强度减弱有关[23, 37]。黄土剖面的牙釉质碳同位素研究则揭示出,中国北部C4植被空间分布模式的演变主要和夏季降水南北移动有关,在中新世-上新世暖期夏季风强盛,森林-草原-沙漠的位置相对现代环境北移;4~2Ma期间,冬季风加强,C4植被发生南移,2Ma以后“最大C4生物量”位于黄土高原地区南部的南侧[28]。临夏盆地在第四纪才有C4植被的出现,可能与3~2Ma之前,东亚夏季风较弱,无法带来足够的夏季降水有关[38]。内蒙古地区牙釉质碳同位素记录显示,C4植被在7~4Ma逐渐成为生态系统中重要组成部分,被认为可能是东亚夏季风增强导致的降水季节性增强所致[39]。另外,在中国南海也有利用沉积黑碳来研究华南地区C4植被长时间尺度演化历史的研究,与高纬地区碳同位素记录不同,在4~2Ma存在C4植被扩张的趋势,其演化机制可能与夏季风演化及火活动有关[32, 34]。
因此,晚上新世以来,高、低纬度地区C4植被的演化历史存在差异,其演化机制不仅有全球性的因素(如大气CO2浓度),也可能与区域气候(温度、降水、季节性等)、天然火等因素有关。目前对东亚中高纬度的黄土高原地区、低纬度的华南地区C4植被演化的研究较多[32, 34, 40]。相比之下,由于缺少良好载体,对中国中部、地处中低纬度长江流域的C4植被长时间尺度的演化历史研究极少,不仅各地区主要驱动机制仍具有争议[40~41],而且高低纬度地区之间演化的差异性也没有得到更多相关的研究。对比研究以上这3个不同纬度带的C4植被演化差异,归纳总结其时空演化规律,可望对长时间尺度中国不同纬度地区的C4植被演化的机制有新的认识。另一方面,土壤碳酸盐碳同位素重建的C3/C4植被的可靠性具有一定争议,而有机质碳同位素重建的演化历史有限,动物牙釉质化石的时间分辨率则很低[40, 42~43],由此,需要更高分辨率的可靠指标来重建长时间尺度的C3/C4植被演化历史。本研究中,我们以南黄海CSDP-1孔贯穿第四纪地层的长岩芯为研究对象,提取沉积物中的黑碳,分析其含量、通量及碳同位素组成(δ13CBC),进而研究中国东部地区自晚上新世以来C3/C4植被演化历史;通过对比分析黄土高原、长江流域、华南地区这3个纬度带自晚上新世以来C4植被演化历史,从时空演化角度分析影响植被演化的机制。
1 研究材料与方法 1.1 研究材料本次研究材料为中国“大陆架钻探计划”(Continental Shelf Drilling Program)于2013年6月在南黄海西部钻取的长柱状沉积物岩芯CSDP-1(如图 1)。CSDP-1孔(34°18′N,122°22′E;水深52.5m)由上海海洋石油局第一海洋地质调查大队“勘407”号海洋工程地质综合调查船钻探取得,岩芯长300.1m,泥质沉积物为主的岩芯平均取芯率大于85 %,砂层的取芯率为60 % ~85 % [4]。

|
图 1 东亚海-陆分布及研究站位图 其中蓝线为河流,黑线为-50m等深线;红色圆点为本次研究所用岩芯CSDP-1孔,白色圆点为本次研究中使用的中国河流表层样品取样点,黄色圆点为本文中引用的站位(包括南海ODP1148站、日本海IODP U1430站、黄土高原灵台剖面和蓝田剖面) Fig. 1 Land-sea distribution of East Asia and locations of the studied sites. Also shown the major rivers in East Asia(blue lines), -50m isobath(black line), the studied core CSDP-1(red dot), river sediment sample sites(white dots)and sites cited in this study(yellow dots)(including ODP Site 1148 in the South China Sea, IODP Site U1430 in the Japan Sea, Lingtai and Lantian sections in the Chinese Loess Plateau) |
岩芯岩性总体可分为三段[4, 6]:1)0~83.00m,以深灰色的粘土质粉砂、粉砂质粘土为主,夹杂少量薄的砂层,含少量碳质斑点,海相化石较多,生物扰动作用较强,总体反映了浅海相沉积环境;2)83.00~148.64m,总体以灰黄色的细砂-中砂为主,夹杂3个粒度较细的层位,以灰色的粘土质粉砂为主,具有少量海相化石,含有锈斑,沉积环境总体以陆相河流沉积为主;3)148.64~300.10m,具有下砂层和上泥层组成的粒度向上变细的多个沉积旋回特征,总体以灰黄色的细砂-中砂为主,夹少量砾石薄层,交错层理发育,叠覆在砂层上的褐黄色或深灰色粘土质粉砂和泥层含大量的锈斑和钙质结核,不含海相化石,泥层含少量陆相化石,生物扰动作用中等,总体反映了陆相河流沉积环境。
根据高分辨率的磁性地层和放射性碳十四(AMS14C)年代学结果,沉积地层相对连续,岩芯底界年龄约3.5Ma[44]。第四系底界(G/M,约2.58Ma)位于约227.16m;松山-布容界线(M/B,约0.78Ma)位于约73.68m[44]。岩芯的年代框架基于古地磁和6个AMS14C测年数据,平均沉积速率约9.5cm/ka[6]。
本研究选取了150个CSDP-1孔样品,以约2m间隔取样,单个样品厚度为2cm。各样品的年代据年代控制点线性内插获得。另外,还选取了11个中国各大河流(乌龙江、闽江、瓯江、钱塘江、长江、淮河、黄河)下游近河口的表层沉积物样品(如图 1),提取黑碳以对比分析。
1.2 研究方法 1.2.1 黑碳的提取与测试分析现有的黑碳测定方法主要包括热氧化法、化学氧化法、分子标志物法和光学法。前两种测定方法主要依据黑碳的热稳定性和化学稳定性,分子标志物法主要测定黑碳中特定的芳香簇组分,光学法则主要根据黑碳组分的光学特性来评估黑碳特性[45]。本次研究采用Lim和Cachier[46]提出的化学氧化法提取黑碳。化学氧化法主要根据黑碳与非黑碳有机质的化学稳定性不同,用一定氧化强度的化学试剂去除环境样品中的非黑碳组分[45];目前该方法已广泛应用于黄土、湖泊、海洋沉积物中黑碳的提取与分析[10, 22, 47~49]。
具体实验方法如下:取约1.5g干样;加入3mol/L的HCl溶液常温反应24 h,去除无机碳酸盐;加入10mol/L的氢氟酸(HF)和1mol/L的盐酸(HCl)混合溶液常温反应24 h,去除硅酸盐矿物;加入10mol/L的HCl溶液常温反应24 h,去除残留的Fe2O3、Al2O3及其他氧化物;加入0.1mol/L的K2Cr2O7和2mol/L的H2SO4混合溶液,55℃水浴加热60 h,去除非黑碳有机质组分。剩余的黑色物质即为黑碳,将其烘干称重。
黑碳碳同位素和含量测试,在中国科学院烟台海岸带研究所分别用稳定同位素质谱仪(Thermo Fisher MAT 253 Plus)和元素分析仪(Flash EA 2000型)完成。其中,碳同位素组成(δ13C)采用相对于国际标准样品(VPDB)表示,对已知碳同位素组成的标样(IAEA-600,δ13C=-27.771 ‰)重复测试显示,测试误差小于±0.2 ‰。黑碳碳元素含量的平行样测试结果显示,相对误差小于5 %。黑碳通量用以下公式计算得到:
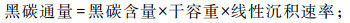
|
黑碳含量指全样中黑碳所占质量百分比。干容重数据来自Zhang等(2019)[6],线性沉积速率数据来自Liu等(2016)[44]。
1.2.2 扫描电镜及能谱分析取少量提取的黑碳用酒精浸泡,用超声波分散,然后吸取适量黑碳悬浮液均匀涂抹在载玻片上,待晾干后用镀膜机在其表面喷金。在青岛海检集团用ISM-7610F型扫描电子显微镜(SEM)观察黑碳形态、大小,用UltraDry能谱仪(EDS)对黑碳元素组成进行能谱分析。
2 结果 2.1 黑碳含量、通量及碳同位素组成CSDP-1孔黑碳的含量和通量(MAR)在3.5Ma以来变化很相似(如图 2),总体呈波动幅度增加的趋势。3.5~0.8Ma,黑碳含量呈波动变化,阶段性增大,最小值为0.039 ‰,最大值为1.86 ‰,平均值为0.331 ‰;黑碳通量整体呈低值,最小值为0.8mg/cm2/ka,最大值为41.3mg/cm2/ka,平均值为7.3mg/cm2/ka。约0.8Ma以来,黑碳含量增大,变化幅度也明显增大,最小值为0.015 ‰,最大值为1.977 ‰,平均值为0.732 ‰;黑碳通量及其变化幅度也相对增大,最小值为0.3mg/cm2/ka,最大值为52.7mg/cm2/ka,平均值为12.8 mg/cm2/ka。

|
图 2 晚上新世以来CSDP-1孔岩性、黑碳含量、通量及δ13CBC变化 岩性特征改自Liu等(2018)[4],SC:粉砂质粘土,CSi:粘土质粉砂,S:粉砂,FS:细砂,MS:中砂,CS:粗砂,G:砾石 Fig. 2 Variation of content, MAR and δ13CBC of black carbon of core CSDP-1 in the South Yellow Sea since the Late Pliocene. The characteristic of the core lithology was simplified from Liu et al. (2018)[4], SC: silty clay, CSi: clayer silt, S: silt, FS: fine sand, MS: medium sand, CS: coarse sand, G: gravel |
CSDP-1孔黑碳δ13C值在3.5Ma以来呈先正偏后负偏的趋势,其变化大致可分为3个阶段:3.5~2.6Ma,δ13C值相对偏负,变化幅度较大(-25.6 ‰至-18.0 ‰),平均值为-21.4 ‰;2.6~0.8Ma,δ13C值整体相对偏正(-23.3 ‰至-17.1 ‰),平均值为-19.8 ‰;约0.8Ma以来,δ13C值相对偏负(-24.3 ‰至-19.1 ‰),平均值为-21.7 ‰。
另外此次研究还选取了从南到北11个中国河流表层沉积物样品提取黑碳分析,结果如表 1所示,长江沉积黑碳的δ13C值最为偏正,其次是淮河,再次是黄河;总体而言,长江、淮河和黄河数据要较其以南河流的数据更为偏正。
| 表 1 中国各河流表层沉积物中黑碳的δ13C值 Table 1 The δ13C values of black carbon of surface sediments of rivers in China |
我们对CSDP-1孔4个层位的黑碳样品做镜下观察,各层位黑碳形态、大小相似,其中具代表性的镜下照片如图 3所示。黑碳颗粒边缘普遍具有棱角,磨圆度中等,分选差-中等。有些黑碳颗粒还保留了原始燃烧植被碎屑的形态特征(如图 3b)。黑碳粒度变化较大,多在几微米-几十微米之间,极少数大的黑碳颗粒粒径可达100μm。黑碳形态多呈板状、片状,粒度小的黑碳磨圆度比大颗粒黑碳相对更好。X射线能谱分析显示,黑碳成分以碳元素为主(图 3)。

|
图 3 CSDP-1孔黑碳扫描电镜照片和能谱结果 cts:计数,keV:千电子伏特;(a)和(c)样品深度约126m,(b)和(d)样品深度约48m Fig. 3 SEM images and EDS results for the black carbon of core CSDP-1. cts: counts, keV: kiloelectron volt; (a)and (c) from the sample at depth of 126m; (b)and (d) from the sample at depth of 48m |
解读沉积黑碳所记录的古环境信息之前,需要首先约束黑碳的来源。通常在远离大陆的深海,风尘是黑碳的主要输送方式。在第四纪干旱化总体增强的趋势下,现代输入西太平洋的粉尘通量约为70 Mt/a[50]。在北太平洋深海沉积物中发现了自早新生代以来的大量木炭,在日本海、东太平洋沉积物中也存在由风力搬运沉积的黑碳;黑碳(木炭)通量一般在0.1~2.0mg/cm2/ka范围内[22, 27, 51];相比之下,在东亚边缘海,如渤、黄、东海,其沉积物主要来自东亚大河如长江和黄河的输入。现代黄河汇入渤海的输沙量约为1100 Mt/a;长江汇入东海的输沙量约为470 Mt/a;其他小河流,如淮河、海河、滦河、辽河等,输沙量相对较低,合计约76 Mt/a;朝鲜半岛的小河流输沙量更低,约18 Mt/a[2]。显然,长江、黄河巨大的输沙量,成为了中国东部边缘海主要的陆源物质包括黑碳的供应者。有研究估算现代长江和黄河分别输送2.0×1011g和1.2×1010g黑碳进入东海和渤海[13]。中国陆架海沉积黑碳含量,渤海湾最高(平均约2.18 ‰),渤海较高(平均约0.84 ‰),黄海尤其南黄海相对较低(平均约0.52 ‰),随着离河口距离的增加,黑碳含量降低,暗示着河流在近海输送黑碳中的重要作用[52~55];现代渤海黑碳通量约30mg/cm2/ka[56]。CSDP-1孔沉积黑碳含量、通量变化幅度大,与岩性变化有着密切关系,黑碳更集中于细粒级碎屑物质中[10]。总的来说,CSDP-1孔黑碳通量在10~20mg/cm2/ka之间,主要由河流输入。
有关南黄海地区沉积物物源的研究多集中于晚第四纪时期[5, 57~58]。前人通过粘土矿物、轻重矿物、元素和同位素地球化学、层序地层等方法,对南黄海不同区域进行了研究,发现南黄海西部及中部泥质区沉积物主要来自长江和黄河,东部有来自朝鲜半岛的物质[3, 57, 59~61];相比之下,对于更长时间尺度如中更新世之前的南黄海沉积物物源的研究相对较少。CSDP-1孔沉积物中碎屑锆石的U-Pb年龄分析表明,至少在3.2Ma以来研究区物源以长江物质为主,0.8Ma以来以黄河物质为主,其中2.2Ma开始可能受到了淮河的影响,但其贡献较低[62]。该岩芯沉积物的粘土矿物、Sr-Nd同位素地球化学分析也揭示,其物源在0.8Ma左右发生了较大的变化,从以长江源为主转变为以黄河源为主;同时,由于中国东部地区的沉降,长江的主河道也由南黄海迁移到了现代长江三角洲地区[6]。物源转换和区域性海侵时间上的一致性表明构造作用是影响黄河和长江演化以及渤海和黄海沉积环境变化的一级控制因素[6]。
黑碳的形态特征和碳同位素组成也是判断其来源的重要标志[22]。我们对CSDP-1孔的黑碳样品做镜下观察,发现黑碳粒径多在数微米-数十微米之间,极少数黑碳粒径可达100μm;边缘普遍具有棱角,有些黑碳还保留了植物结构形态特征(图 3)。黑碳的形态特征表明,黑碳由河流输入为主,风力输入很少。长江流域、黄河流域气候差异大,覆盖植被不同[63],因此,长江、黄河沉积物黑碳的碳同位素组成应当存在差异。黄土高原有机质δ13C显示约0.8Ma与表层δ13C值相近[23, 33],但是由于缺乏长时间尺度的长江流域C3/C4植被演化记录,难以将地质历史时期的两个流域植被进行对比,仅从孢粉分析,约0.8Ma与现代长江流域植被没有发生明显变化[64~65]。本研究中,长江和黄河沉积物的黑碳碳同位素组成平均值分别为-20.2 ‰和-21.6 ‰ (见表 1),长江相对黄河更偏正。在0.8Ma左右黑碳δ13C明显负偏,结合上述碎屑态物源研究表明,黑碳物源在中更新世后由长江源变为黄河源。
3.2 黑碳碳同位素组成含义C3、C4植物由于光合作用途径不同,具有明显不同的δ13C值范围[19]。这种特征在植被不完全燃烧形成的黑碳中保留了下来。但值得注意的是,在植被燃烧过程中,碳同位素可能会发生分馏,即燃烧不完全产物的碳同位素组成与原植物的碳同位素组成有所差异[66]。C3、C4植物燃烧实验结果对比表明,C3植物燃烧形成的黑碳与原植物的碳同位素差值变化范围在-3 ‰到+3 ‰之间,平均约为-0.3 ‰,C4植物燃烧形成的黑碳与原植物的碳同位素差值变化范围在-10 ‰到+3 ‰之间,平均约为-1.7 ‰ [48]。碳同位素的分馏除了与植被种类有关,还与燃烧时的温度有关[66~67]。利用化学氧化法提取的具有稳定结构的黑碳,其形成的温度条件相对较高(400~600℃)[45, 66],因此燃烧温度对化学氧化法提取的黑碳碳同位素分馏效应的影响较小。另外,地质历史时期的大气CO2浓度及δ13C值变化对C3、C4植物本身的δ13C值也有影响[68~69]。有研究基于大气CO2浓度变化可以控制进入植物的13C数量,发现C3植物的碳同位素组成与大气CO2的δ13C值之间的差值和大气CO2浓度之间存在显著的双曲线函数关系[68]。但自晚上新世以来这种分馏效应是非常有限的,可以忽略不计,且自晚上新世以来,大气CO2的δ13C值变化也很小(<0.5 ‰)[69],因此同样可忽略其对植物本身碳同位素组成的影响。
此外,研究表明黄土高原表层土壤中黑碳的碳同位素组成(δ13CBC)与土壤有机质的碳同位素组成(δ13CSOC)具有显著的相关关系,δ13CBC与δ13CSOC的平均差值不超过1.5 ‰,且黄土高原表层土壤的δ13CBC值空间变化与该区域植被的空间分布变化是一致的[70]。因此,δ13CBC值常被作为重建源区古植被的有效指标。在黄土和海洋沉积中,具有高度稳定性的黑碳已经被有效应用于C4植被演化的重建[22, 27, 49]。
当确定了各地质历史时期C3、C4植被的碳同位素组成端元,则可根据下面的公式估算得到C4植被在燃烧植被生物量中所占百分比:
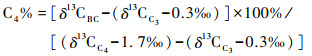
|
其中,C4 % 为C4植被在燃烧植被生物量中所占百分比,δ13CBC为黑碳碳同位素组成,δ13CC4和δ13CC3分别为C4植被碳同位素组成端元和C3植被碳同位素组成端元。本次研究,只利用δ13CBC值作为C3/C4植被燃烧生物量变化的指标,不估算C4植被对燃烧生物量的贡献。CSDP-1孔沉积黑碳δ13C值在3.5Ma以来发生了两次明显的变化(图 2):在约2.6Ma显著正偏,指示C4植被相对丰度增加,对燃烧植被生物量的贡献增多;约1.0Ma以来,δ13C值开始负偏,尤其在约0.8Ma明显负偏,指示了C3植被的相对丰度增大。
3.3 全球C3/C4植被分布与演化 3.3.1 现代C3、C4植被分布现代C4植物分布与气候因子(温度、降水、季节性和海拔等)之间的关系非常复杂。世界范围内C4植物种属的地理分布的系统总结发现,C4植物多分布在中低纬地区和低海拔地区,尤其在热带炎热、开放的环境中,C4植物具有绝对优势[71]。北美洲各地区C4草种属丰度与日照量、降水、温度等气候记录相关性分析显示,植物生长季最低温与C4草种属丰度相关性最高,因此,生长季的低温不利于C4草生长[72]。海拔高度对C3、C4植被分布也有影响。阿根廷地区不同海拔的C3、C4草分布研究发现,在低海拔地区,C4草的种属和植被覆盖均占优势,而高海拔地区C3草占优势,因此研究人员认为C3、C4植物分布与蒸发和温度高度相关,与降水相关性较低[73]。但是,也有研究认为C4植被的分布与水分条件有密切关系[74~75]。澳大利亚地区禾本科中的C3、C4植物种属、分布与温度、降水的相关性分析表明,在各自适合的温度条件下,C3、C4种属均随着降水的增加而增多;在炎热潮湿的夏季,C4种达最大值,在凉爽潮湿的春季,C3种最多;而C4种属数随气温的下降和/或夏季雨量的减少而减少,C3种属数随气温的升高和/或春季降雨量的减少而减少[74]。中国C4植物的地理分布显示,C4植物种数(尤其是禾本科和莎草科)随着大气温度与水分增高而增加,但藜科的C4植物种数随着大气温度与水分增高而减少[75]。这说明不同种的C4植物可能适应不同的生境,需要在未来的C4植被演化的研究中考虑这一因素。表土总有机质碳同位素数据估算的环北太平洋地区(中国东部地区、北美大平原、澳大利亚地区)现代植被中C3和C4植物的相对生物量贡献比例的对比表明[21],温度条件对C4植物生物量具有决定性影响。温度过低的区域,如中国东部地区年均温度如果低于12℃左右,无论降水量怎么变化,C4植物相对丰度都极为有限;而且,当温度条件满足了C4植物生长的需要,随着温度进一步上升,降水量应相应地减少,C4植物才能保持其优势地位[21]。
本研究选取了从南到北的11个中国河流表层沉积物样品开展了黑碳分析,结果如表 1所示。华南地区黑碳δ13C值在-25 ‰至-24 ‰之间,中纬度地区黑碳δ13C值在-22 ‰至-20 ‰之间,中纬度地区具有相对更偏正的δ13C值,C4植物的相对含量更高。同样地,中国东部表土总有机质和长链正构烷烃碳同位素以及表土植硅体碳同位素结果显示[76~77],30°~40°N范围内C4植物相对丰度更高,气候条件更适宜C4植物生长。相比之下,华南地区降水量、温度更高,降水的季节也更宽泛,在春季的高降水有利于C3植物生长,占据了生态位,而不利C4植物生长;中纬度地区随着降水量的降低同时降水也更集中于炎热的夏季,C4植被生长优势凸显,具有更高的相对含量[78~79]。
3.3.2 冰期-间冰期尺度C3/C4植被演化对全球各纬度带C3/C4植被在冰期-间冰期尺度的演化趋势进行综合对比,可以发现,在低纬度地区,从末次冰期到全新世,C4植被多被C3植被取代;而中高纬度地区,从末次冰期到全新世,C4植被具有普遍扩张的趋势[41]。例如中国东南部湖泊沉积、南海海相沉积、热带哥伦比亚湖泊沉积、孟加拉湾海相沉积中的正构烷烃δ13C值都记录到C4植被在间冰期(全新世)相对减少,C3乔木相对增多;而中国黄土高原和东北黄土沉积中有机碳δ13C值、日本海海相沉积中正构烷烃δ13C值则记录到C4植被在全新世发生相对扩张[80~87]。由此可见,不同纬度带的C3、C4植被在冰期-间冰期气候条件发生转变时具有不同的竞争优势。参考现代植被的分布特征,在降水量丰富的低纬度地区,C3乔木具有更大的竞争优势,当冰期来临,大气CO2浓度大幅度降低,降水量减少,气候更为干旱,同时,温度下降幅度很小[85, 88~89],C4植物的竞争优势相比C3植物增强,C4生物量随之增加;而中高纬度地区,冰期来临时,降水量降低,但同时温度的下降幅度也很大[90],难以满足C4植物的生长;因此间冰期时期,温度升高、暖季降水量增加,C4植物的竞争优势才开始凸显[41, 91~92]。
3.3.3 晚上新世以来C3/C4植被演化长时间尺度C3/C4植被演化研究显示,全球多个区域均发现了显著的C4植被扩张现象,而这些C4植被的扩张事件主要集中出现在晚中新世时期(8~6Ma),被认为C4植被扩张具有全球同步性[19, 26, 28~29]。但在后来的研究发现,C4植被的扩张并不完全具有全球同步性[22, 24, 93],因此,长时间尺度的C3/C4植被演化也与区域气候密切相关。然而,通过对比分析全球范围内C4植被的演化趋势,同样可以发现植被的长期演化具有纬度差异特征[22~25, 27, 29, 32, 94~95](图 4):在中低纬度地区,自第四纪以来,C4植被多呈相对增加的趋势;在中高纬度地区则呈下降的趋势。例如北美(<37°N)、北非和巴基斯坦的土壤碳酸盐δ13C值、北非和澳大利亚的叶蜡正构烷烃δ13C值以及南海和南黄海(本研究)沉积黑碳δ13C值均记录到在晚上新世-早更新世C4植被的相对扩张趋势;而黄土高原碳酸盐和土壤有机碳δ13C值和东太平洋黑碳δ13C值显示了C4植被的相对减少。C4植被演化的纬度差异可能意味着区域的气候因素如温度起到了重要作用。如前文所述,降水和温度的绝对值变化控制着植被的分布与演化。在中低纬度地区,第四纪两极冰盖扩张,干旱进一步增强[96],C4植被的生存优势可能超过了C3植被,导致了中低纬度地区C4植被的扩张;而在中高纬度地区,第四纪开始气候变得更为干冷[97],温度的降低可能成为了C4植被生长的限制因素[22, 27]。

|
图 4 晚上新世以来CSDP-1孔记录的C3/C4植被演化及全球对比 碳同位素记录来自南黄海地区(本研究)、北美[95]、东非[25, 94]、巴基斯坦北部[29]、澳大利亚[24]、南海[32]、黄土高原[23]以及东太平洋[27] Fig. 4 Evolution of C3/C4 vegetation recorded by carbon isotope data at core CSDP-1 and other sites of the globe since the Late Pliocene. Carbon isotope data from the South Yellow Sea(this study), the Great Plains[95], Eastern Africa[25, 94], Northern Pakistan[29], Australia[24], the South China Sea[32], the Loess Plateau[23]and the northeastern equatorial Pacific[27] |
黑碳物源结果显示,3.5~0.8Ma,CSDP-1孔沉积黑碳主要由长江输入。位于中低纬度的长江流域地区在3.0~2.6Ma发生C4植被相对丰度的增加,与华南地区C3/C4植被的演化趋势类似[32]。C3/C4植被演化的驱动机制复杂,可能与气候、大气CO2以及火等因素有关[22]。
火在C4植被取代C3植被的过程中可能会起到重要作用[27]。当C3乔木被大量燃烧后,会形成一个开放的生态环境,C4植物能够优先适应并生长,占据生态位,因此有研究认为火是导致C4植被扩张的一个重要因素[27, 31, 34, 98~99]。本研究中的沉积黑碳是植被不完全燃烧的产物,火燃烧的植被越多,形成的黑碳也越多,从而黑碳含量和通量可以反映源区火活动的频率或强度变化[10, 22]。CSDP-1孔黑碳含量和通量在晚上新世以来呈阶段性的波动变化(图 5),在约2.6Ma北半球冰盖扩张以来并没有明显的增强,而约0.8Ma以来则波动幅度明显增大,后者可能与中更新世转型之后全球气候冰期-间冰期波动幅度增加[97]引起黄河流域火活动变率增强有关;也可能由于此后黄海开始大规模海侵,沉积物输送受到海平面波动影响[4, 6]。无论如何,黑碳含量和通量的长期趋势明显不同于其碳同位素组成反映的植被类型变化(图 5),后者呈现三段式波动,因此,可以认为中国东部地区火与植被之间在百万年时间尺度上的相关性并不显著,火不是该区域驱动植被长期演化的重要因素。但是,我们也注意到CSDP-1孔黑碳含量和通量的几个万年时间尺度的特征高值,即火活动峰值,对应了黑碳δ13C负偏峰值(图 2),在东太平洋长时间记录和中国北方岱海湖短时间记录中也有类似记录[27, 48],可能是自然为平衡生态系统生物量引起的强野火事件清除了源区的森林植被,暗示火活动可能在短时间尺度上对植被演化有较重要影响。

|
图 5 晚上新世以来CSDP-1孔黑碳记录及C4植被扩张驱动因子对比 驱动因子包括全球变冷(全球深海氧同位素[97])、火活动(黑碳含量、通量)、干旱度(柴达木盆地孢粉指标[100]、长江三角洲平原孢粉指标[65]、黄土高原风尘通量[101]、北太平洋风尘通量[102])、夏季风演化[103]以及大气CO2浓度[104];青藏高原阶段性隆升与干旱演化作对比[6, 105~106] Fig. 5 Potential driving forces of C4 plant expansion since the Late Pliocene. The potential driving forces include global cooling[97], fire activity(black carbon content and MAR), aridity(sporopollen records in the western Qaidam Basin[100], sporopollen records in the Yangtze River Delta[65], eolion deposition on the Loess Plateau[101]and eolion deposition in the north Pacific[102]), East Asian summer monsoon evolution[103]and atmospheric CO2 concentration[104]. For comparison, trend of stepwise uplift of Tibetan Plateau and drying since the Late Pliocene are shown[6, 105~106] |
除了火活动外,夏季风降水强度也是C4植物演化的潜在影响因素[23]。C3、C4植物的生长季有所差异。在凉爽的春季,随着降水、温度的增加,植物开始生长,到了炎热的夏季,高温导致植被的蒸腾作用增强,叶片的气孔在高温条件下闭合,叶片中的CO2浓度降低,导致C3植物的光合作用效率降低,但C4植物仍能维持高的光合作用效率[18, 71, 78]。因此,在炎热的夏季C4植物具有较大的生存优势[18]。生长季降水的增多有利于植物的生长,因此夏季(暖季)降水的增加,有利于C4植物的扩张[79]。根据黄土高原地区黄土-古土壤剖面磁化率、碳酸盐含量重建的夏季风指标,自晚上新世以来,夏季风强度先减弱后增强,在约2.7Ma以来,夏季降水开始减少[103],而C4植物反而开始扩张(图 5)。因此,夏季降水的变化不是影响3.0~2.6Ma中国长江流域C4植被相对丰度增加的主要因素。
当温度满足C4植物生长的条件,高的降水量有利于C3乔木的生长,随着降水量的降低,干旱程度的增强,C4植物的生长优势逐渐凸显[21]。如图 5,柴达木地区蒿/藜科(A/C)比值、长江流域乔木类孢粉/非乔木类(AP/NAP)比值、黄土高原及北太平洋粉尘记录等干旱指标显示自晚上新世以来,亚洲内陆干旱程度逐渐增强,尤其在约2.6Ma干旱程度显著加剧[65, 100~102],对比CSDP-1孔黑碳δ13C变化,约2.6Ma长江流域C4植被相对丰度的增加很可能受到了干旱因素的影响。2.6Ma以后黄土-古土壤剖面磁化率显示东亚夏季风开始减弱,降水总体减少,变化幅度增大,而黄土粗粒级含量、Al通量以及气候模拟结果显示东亚冬季风持续增强,意味着中亚干旱的加剧[96, 107~108],且东亚夏季风气候变化主要响应北半球夏季太阳辐射变化[109]。3.5Ma以来,深海氧同位素曲线(δ18O值)显示出明显正偏的趋势,北半球高纬地区冰盖形成并扩张,全球进入两极有冰盖时期,导致全球气候变冷,大气中的水汽含量降低[97, 110~111]。新生代以来青藏高原的隆升也被认为对亚洲季风增强和干旱的加剧具有重要作用,通过改变大气环流模式并阻挡来自南部的水汽从而对亚洲气候产生影响[96]。青藏高原在晚上新世以来有几次阶段性的快速隆升事件:约3.6Ma的青藏高原运动A幕、2.6Ma的青藏高原运动B幕、1.8~1.7Ma的青藏高原运动C幕、1.2~0.6Ma的昆黄运动和0.15Ma的共和运动,高原隆升的雨影效应可能加剧了亚洲干旱[106]。东亚冬季风与夏季风在7.0~3.2Ma以前正相关,在2.7Ma以后转变为负相关关系,驱动机制可能由太阳辐射驱动转变为青藏高原的阶段性隆起和北半球冰盖发育的综合效应影响,气候变得更加干冷[103]。因此,约2.6Ma东亚干旱度的增强可能是青藏高原隆升和北半球冰盖扩张共同作用的结果,该过程减弱了海洋水汽的蒸发和水汽向亚洲内陆的输送,从而加剧了亚洲大陆的干旱化。如前文所述,全球中低纬度地区在干旱化持续增强的条件下,C4植被具有逐渐扩张的趋势[21]。相对于第四纪,晚上新世时期,长江流域降水充沛,利于C3乔木生长[65];约2.6Ma干旱的增强有利于C4植物获得更高的竞争优势。
此外,大气CO2浓度也是控制C4植被演化的重要因素[19, 112~115]。大气中CO2浓度的下降被认为是驱动晚中新世时期C4植被扩张的一个关键因素[19]。当大气CO2浓度降低,C3植物由于其高的CO2补偿点,光合作用效率明显下降,而C4植物CO2补偿点很低,仍能保持高的光合作用效率[17~18, 116~117]。在晚上新世-早更新世,大气CO2浓度显著下降,由350 ppm下降到250 ppm[104](图 5),因此,这也有可能是C4植被相对丰度增加的原因之一。综上,我们认为3.0~2.6Ma长江流域C4植被相对丰度的增加可能是大气CO2浓度降低和区域干旱度增强叠加的结果。
然而,自约1.0~0.8Ma开始,南黄海CSDP-1孔黑碳碳同位素指示的C4植被相对丰度呈降低趋势(图 5)。尽管物源研究结果显示,在约0.8Ma之后CSDP-1孔的主要物质来源发生了变化,由古长江变为了古黄河[6, 62],源区植被可能存在差异,影响了约0.8Ma黑碳δ13C的负偏。但是,有意思的是,源于中亚地区被搬运至日本海、东太平洋的风尘黑碳,其δ13C值在约1.2~0.8Ma开始均有负偏的趋势[22, 27],南海北部ODP1148站黑碳δ13C值在约0.8Ma开始也有类似负偏趋势[32](图 6),而这和上述干旱指标均指示亚洲内陆干旱化持续增强的趋势正好相反(图 5),这暗示着可能有另一因素逐渐成为中更新世之后控制东亚C4植被演化的首要机制。

|
图 6 晚上新世以来东亚-太平洋地区黑碳记录及海陆温度对比 黑碳记录包括南黄海(本研究)、日本海[22]、东太平洋[27]以及南海[32];古温度记录包括黄土高原陆表温度[90]、南海海表温度[119]、西北太平洋海表温度[120]以及深海氧同位素[97]右侧,绿色和黄色阴影分别代表以黄河和长江为主的物源 Fig. 6 Black carbon record from East Asia-Pacific regions and land-sea temperature reconstructions since the Late Pliocene. Black carbon records include South Yellow Sea(this study), Japan Sea[22], eastern Pacific[27]and South China Sea[32]. Paleotemperature records include land surface temperature (LST) on the Loess Plateau[90], sea surface temperature(SST)of the South China Sea[119], SST of the northwest Pacific[120]and the deep sea oxygen isotope[97]. The green and yellow shades in right represent the provenance dominated by the Yellow and Yangtze Rivers, respectively |
前文提到,在中高纬度地区温度是限制C4植被生长的重要因素,在温度过低的条件下,无论降水量怎么变化,C4植物相对丰度都极为有限[21]。而且,沉积记录由南海-南黄海-日本海-东北太平洋,其物质来源由华南地区-长江(黄河)流域-中亚地区,纬度越高,陆地温度受到全球温度降低的影响越显著[97],植被对温度变化的响应越早。1.2~0.6Ma时期为中更新世气候转型期(MPT),全球气候变化的主导周期从早更新世时期高频低幅的4万年周期转变为中更新世之后低频高幅的10万年周期[97, 118]。MPT后伴随着全球冰量大幅度增加的全球性气候变冷,东亚冬季风、夏季风增强,出现极端气候期[119~122]。由GDGTs重建的80万年以来黄土高原地区陆表古温度结果显示,冰期-间冰期之间存在4~10℃的差异[90](图 6),而现代C4植物占优势的地区年均温应在12~16℃之间[21],寒冷的冰期气候显然不适合C4植被生长。即使暖湿的间冰期,C4具有较高丰度[85],但在长时间尺度上,温度还是影响了C4植被的生长,进而影响了碳同位素的记录[123]。因此,从约1.0~0.8Ma开始,CSDP-1孔黑碳δ13C值的负偏、C4植被相对丰度的减少可能是由于北半球冰盖扩展及逐渐降低的温度成为了限制冰期C4植被生长的重要因素,再加上物质源区的改变,北方植被更容易受到温度的影响。
值得注意的是,MPT之后,温度在千年尺度上大幅波动,总体呈降低的趋势[90],但在约0.4Ma,δ13CBC值有一个小幅的正偏(图 5)。同样地,黄土高原有机质δ13C也显示了约0.4Ma的正偏[23, 33]。冰期过低的温度限制了C4植物生长,但在温暖湿润的间冰期,C4植被大幅扩张[85]。尤其在MPT后,夏季风明显增强,带来更多夏季降水[103],更有利于间冰期C4植物的生长。因此,夏季风的增强可能导致了这一阶段C4植被相对丰度的增加。
4 结论本文通过南黄海大陆架科学钻探CSDP-1孔沉积黑碳的含量、通量和碳同位素组成分析,并结合现代C4植被分布及不同纬度地区C3/C4植被演化历史研究,主要取得了以下认识:
(1) 从现代-构造时间尺度上,C3/C4植被演化趋势均有纬度差异。中低纬度地区干旱化可能更有利于C4植被相对丰度的增加,而中高纬度地区更容易受到低温的限制。
(2) 黑碳SEM-EDS、碳同位素组成并结合以前碎屑物源研究,结果表明南黄海CSDP-1孔沉积黑碳主要来自长江和黄河。3.5~0.8Ma之间,沉积物源主要来自古长江,自约0.8Ma始,物源由古长江变为古黄河。
(3) 晚上新世(3.0~2.6Ma),CSDP-1孔黑碳同位素(δ13CBC)值正偏,暗示源区(长江流域)C4植被相对丰度增加,综合不同纬度地区记录推测干旱化加强可能对中低纬度地区C4植被演化有重要影响,同时,大气CO2浓度降低也可能是C4植被扩张的原因之一。
(4) 自中更新世(约1.0Ma)以来,研究岩芯δ13CBC值整体偏负,表明源区C4植被相对含量减少;综合东亚区域性的碳同位素负偏记录推测,全球气候变冷及源区的北移,使得温度成为了当时影响C4植被演化的重要因素。
致谢: 感谢黄海大陆架科学钻探的全体科学家和船员的辛苦努力;感谢审稿专家和杨美芳老师对初稿提出的许多宝贵意见;感谢中国科学院烟台海岸带研究所谭扬老师在实验测试中给予的帮助。
| [1] |
秦蕴珊, 赵一阳, 陈丽蓉. 黄海地质[M]. 北京: 海洋出版社, 1989: 1-289. Qin Yunshan, Zhao Yiyang, Chen Lirong. Geology of the Yellow Sea[M]. Beijing: China Ocean Press, 1989: 1-289. |
| [2] |
Milliman J D, Farnsworth K L. River Discharge to the Coastal Ocean: A Global Synthesis[M]. Cambridge: Cambridge University Press, 2011: 1-394.
|
| [3] |
蓝先洪, 张宪军, 刘新波, 等. 南黄海表层沉积物粘土矿物分布及物源[J]. 海洋地质与第四纪地质, 2011, 31(3): 11-16. Lan Xianhong, Zhang Xianjun, Liu Xinbo, et al. Distribution pattern of clay minerals in surface sediments of South Yellow Sea and their provenance[J]. Marine Geology & Quaternary Geology, 2011, 31(3): 11-16. |
| [4] |
Liu J, Zhang X, Mei X, et al. The sedimentary succession of the last~3.50 Myr in the western South Yellow Sea: Paleoenvironmental and tectonic implications[J]. Marine Geology, 2018, 399: 47-65. DOI:10.1016/j.margeo.2017.11.005 |
| [5] |
Yang S, Jung H S, Lim D I, et al. A review on the provenance discrimination of sediments in the Yellow Sea[J]. Earth-Science Reviews, 2003, 63(1-2): 93-120. DOI:10.1016/S0012-8252(03)00033-3 |
| [6] |
Zhang J, Wan S, Clift P D, et al. History of Yellow River and Yangtze River delivering sediment to the Yellow Sea since 3.5 Ma: Tectonic or climate forcing?[J]. Quaternary Science Reviews, 2019, 216: 74-88. DOI:10.1016/j.quascirev.2019.06.002 |
| [7] |
Huang J, Wan S, Zhang J, et al. Mineralogical and isotopic evidence for the sediment provenance of the western South Yellow Sea since MIS 3 and implications for paleoenvironmental evolution[J]. Marine Geology, 2019, 414: 103-117. DOI:10.1016/j.margeo.2019.05.011 |
| [8] |
钟福昌, 向荣, 杨艺萍, 等. 黄海中部泥质区底栖有孔虫群落对早全新世海平面快速上升的响应[J]. 第四纪研究, 2020, 40(3): 739-750. Zhong Fuchang, Xiang Rong, Yang Yiping, et al. Rapid sea level rise during the Early Holocene in the central Yellow Sea mud area: Evidence from benthic foraminifera[J]. Quaternary Sciences, 2020, 40(3): 739-750. |
| [9] |
仇建东, 刘健, 张勇, 等. 南黄海西北部晚更新世以来的沉积环境演化[J]. 第四纪研究, 2020, 40(3): 673-689. Qiu Jiandong, Liu Jian, Zhang Yong, et al. Sedimentary environment evolution in the northwestern South Yellow Sea since the Late Pleistocene[J]. Quaternary Sciences, 2020, 40(3): 673-689. |
| [10] |
Pei W, Wan S, Clift P D, et al. Human impact overwhelms long-term climate control of fire in the Yangtze River Basin since 3.0 ka BP[J]. Quaternary Science Reviews, 2020, 230: 106165. DOI:10.1016/j.quascirev.2020.106165 |
| [11] |
Goldberg E D. Black Carbon in the Environment[M]. New York: John Wiley & Sons Ltd., 1985: 1-198.
|
| [12] |
Schmidt M W I, Noack A G. Black carbon in soils and sediments: Analysis, distribution, implications, and current challenges[J]. Global Biogeochemical Cycles, 2000, 14(3): 777-793. DOI:10.1029/1999GB001208 |
| [13] |
Xu C, Xue Y, Qi Y, et al. Quantities and fluxes of dissolved and particulate black carbon in the Changjiang and Huanghe rivers, China[J]. Estuaries and Coasts, 2016, 39(6): 1617-1625. DOI:10.1007/s12237-016-0122-0 |
| [14] |
Brodowski S, John B, Flessa H, et al. Aggregate-occluded black carbon in soil[J]. European Journal of Soil Science, 2006, 57: 539-546. DOI:10.1111/j.1365-2389.2006.00807.x |
| [15] |
张曼, 段阜涛, 李刚, 等. 新疆SCZ17剖面黑碳记录的16-12 ka火灾历史及其驱动因素探究[J]. 第四纪研究, 2020, 40(1): 165-174. Zhang Man, Duan Futao, Li Gang, et al. A study on the fire history and its driving factors during 16-12 ka based on black carbon in SCZ17 section, Xinjiang[J]. Quaternary Sciences, 2020, 40(1): 165-174. |
| [16] |
孙伟伟, 沈吉. 末次间冰期以来兴凯湖黑碳的沉积记录及其环境意义[J]. 第四纪研究, 2020, 40(1): 157-164. Sun Weiwei, Shen Ji. A black carbon record from Lake Xingkai in Northeast China and implication for environmental reconstruction since the last interglacial period[J]. Quaternary Sciences, 2020, 40(1): 157-164. |
| [17] |
贺娟, 汪品先. 晚中新世植被变更与光合作用演化[J]. 地球科学进展, 2005, 20(6): 618-626. He Juan, Wang Pinxian. Vegetation change in Late Miocene and evolution of photosynthesis[J]. Advances in Earth Science, 2005, 20(6): 618-626. DOI:10.3321/j.issn:1001-8166.2005.06.004 |
| [18] |
Sage R F, Sage T L, Kocacinar F. Photorespiration and the evolution of C4 photosynthesis[J]. Annual Review of Plant Biology, 2012, 63(1): 19-47. DOI:10.1146/annurev-arplant-042811-105511 |
| [19] |
Cerling T E, Harris J M, MacFadden B J, et al. Global vegetation change through the Miocene/Pliocene boundary[J]. Nature, 1997, 389(6647): 153-158. DOI:10.1038/38229 |
| [20] |
Edwards E J, Osborne C P, Stromberg C A E, et al. The origins of C4 grasslands: Integrating evolutionary and ecosystem science[J]. Science, 2010, 328(5978): 587-591. DOI:10.1126/science.1177216 |
| [21] |
饶志国, 朱照宇, 贾国东, 等. 环北太平洋地区现代植被中C3/C4植物相对丰度与气候条件关系研究[J]. 科学通报, 2010, 55(12): 1134-1140. Rao Zhiguo, Zhu Zhaoyu, Jia Guodong, et al. Relationship between climatic conditions and the relative abundance of modern C3 and C4 plants in three regions around the North Pacific[J]. Chinese Science Bulletin, 2010, 55(12): 1134-1140. |
| [22] |
Shen X, Wan S, Colin C, et al. Increased seasonality and aridity drove the C4 plant expansion in Central Asia since the Miocene-Pliocene boundary[J]. Earth and Planetary Science Letters, 2018, 502: 74-83. DOI:10.1016/j.epsl.2018.08.056 |
| [23] |
An Z, Huang Y, Liu W, et al. Multiple expansions of C4 plant biomass in East Asia since 7 Ma coupled with strengthened monsoon circulation[J]. Geology, 2005, 33(9): 705-708. DOI:10.1130/G21423.1 |
| [24] |
Andrae J W, McInerney F A, Polissar P J, et al. Initial expansion of C4 vegetation in Australia during the Late Pliocene[J]. Geophysical Research Letters, 2018, 45(10): 4831-4840. DOI:10.1029/2018GL077833 |
| [25] |
Feakins S J, Levin N E, Liddy H M, et al. Northeast African vegetation change over 12 m.y.[J]. Geology, 2013, 41(3): 295-298. DOI:10.1130/G33845.1 |
| [26] |
Huang Y, Clemens S C, Liu W, et al. Large-scale hydrological change drove the Late Miocene C4 plant expansion in the Himalayan foreland and Arabian Peninsula[J]. Geology, 2007, 35(6): 531-534. DOI:10.1130/G23666A.1 |
| [27] |
Kim D, Lee Y I, Hyeong K, et al. Terrestrial biome distribution in the Late Neogene inferred from a black carbon record in the northeastern equatorial Pacific[J]. Scientific Reports, 2016, 6: 32847. DOI:10.1038/srep32847 |
| [28] |
Passey B H, Ayliffe L K, Kaakinen A, et al. Strengthened East Asian summer monsoons during a period of high-latitude warmth? Isotopic evidence from Mio-Pliocene fossil mammals and soil carbonates from Northern China[J]. Earth and Planetary Science Letters, 2009, 277(3-4): 443-452. DOI:10.1016/j.epsl.2008.11.008 |
| [29] |
Quade J, Cerling T E, Bowman J R. Development of Asian monsoon revealed by marked ecological shift during the Latest Miocene in Northern Pakistan[J]. Nature, 1989, 342(6246): 163-166. DOI:10.1038/342163a0 |
| [30] |
Quade J, Cater J M L, Ojha T P, et al. Late Miocene environmental change in Nepal and the Northern Indian subcontinent: Stable isotopic evidence from paleosols[J]. Geological Society of America Bulletin, 1995, 107(12): 1381-1397. DOI:10.1130/0016-7606(1995)107<1381:LMECIN>2.3.CO;2 |
| [31] |
Hoetzel S, Dupont L, Schefuß E, et al. The role of fire in Miocene to Pliocene C4 grassland and ecosystem evolution[J]. Nature Geoscience, 2013, 6(12): 1027-1030. DOI:10.1038/ngeo1984 |
| [32] |
Jia G, Peng P, Zhao Q, et al. Changes in terrestrial ecosystem since 30 Ma in East Asia: Stable isotope evidence from black carbon in the South China Sea[J]. Geology, 2003, 31(12): 1093-1096. DOI:10.1130/G19992.1 |
| [33] |
Sun J, Lü T, Zhang Z, et al. Stepwise expansions of C4 biomass and enhanced seasonal precipitation and regional aridity during the Quaternary on the southern Chinese Loess Plateau[J]. Quaternary Science Reviews, 2012, 34: 57-65. DOI:10.1016/j.quascirev.2011.12.007 |
| [34] |
Zhou B, Bird M, Zheng H, et al. New sedimentary evidence reveals a unique history of C4 biomass in continental East Asia since the Early Miocene[J]. Scientific Reports, 2017, 7: 170. DOI:10.1038/s41598-017-00285-7 |
| [35] |
Ding Z, Yang S. C3/C4 vegetation evolution over the last 7.0 Myr in the Chinese Loess Plateau: Evidence from pedogenic carbonate δ13C[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2000, 160(3-4): 291-299. DOI:10.1016/S0031-0182(00)00076-6 |
| [36] |
杨石岭, 丁仲礼, 顾兆炎, 等. 灵台红粘土-黄土剖面晚中新世以来钙质结核的碳同位素记录及其古植被指示意义[J]. 科学通报, 1998, 43(21): 2323-2326. Yang Shiling, Ding Zhongli, Gu Zhaoyan, et al. Carbon isotopic records of calcareous nodules since Late Miocene in the Lingtai section and their paleo-vegetation implications[J]. Chinese Science Bulletin, 1998, 43(21): 2323-2326. DOI:10.3321/j.issn:0023-074X.1998.21.019 |
| [37] |
Kaakinen A, Sonninen E, Lunkka J P. Stable isotope record in paleosol carbonates from the Chinese Loess Plateau: Implications for Late Neogene paleoclimate and paleovegetation[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2006, 237(2-4): 359-369. DOI:10.1016/j.palaeo.2005.12.011 |
| [38] |
Wang Y, Deng T. A 25 m. y. isotopic record of paleodiet and environmental change from fossil mammals and paleosols from the NE margin of the Tibetan Plateau[J]. Earth and Planetary Science Letters, 2005, 236(1-2): 322-338. DOI:10.1016/j.epsl.2005.05.006 |
| [39] |
Zhang C, Wang Y, Deng T, et al. C4 expansion in the central Inner Mongolia during the Latest Miocene and Early Pliocene[J]. Earth and Planetary Science Letters, 2009, 287(3-4): 311-319. DOI:10.1016/j.epsl.2009.08.025 |
| [40] |
李朝柱, 张晓, 许元斌, 等. 黄土高原地区晚中新世以来陆地植被C3/C4植物相对丰度演化研究进展[J]. 地球科学进展, 2012, 27(3): 284-291. Li Chaozhu, Zhang Xiao, Xu Yuanbin, et al. Reviews on the reconstructed C3/C4 variations since the Late Miocene in the Chinese Loess Plateau[J]. Advances in Earth Science, 2012, 27(3): 284-291. |
| [41] |
饶志国, 陈发虎, 张晓, 等. 末次冰期以来全球陆地植被中C3/C4植物相对丰度时空变化基本特征及其可能的驱动机制[J]. 科学通报, 2012, 57(18): 1633-1645. Rao Zhiguo, Chen Fahu, Zhang Xiao, et al. Spatial and temporal variations of C3/C4 relative abundance in global terrestrial ecosystem since the Last Glacial and its possible driving mechanisms[J]. Chinese Science Bulletin, 2012, 57(18): 1633-1645. |
| [42] |
Rao Z, Zhu Z, Chen F, et al. Does δ13Ccarb of the Chinese loess indicate past C3/C4 abundance? A review of research on stable carbon isotopes of the Chinese loess[J]. Quaternary Science Reviews, 2006, 25(17-18): 2251-2257. DOI:10.1016/j.quascirev.2006.03.013 |
| [43] |
王国安. 稳定碳同位素在第四纪古环境研究中的应用[J]. 第四纪研究, 2003, 23(5): 471-484. Wang Guo'an. Application of stable carbon isotope for paleoenvironmental research[J]. Quaternary Sciences, 2003, 23(5): 471-484. DOI:10.3321/j.issn:1001-7410.2003.05.002 |
| [44] |
Liu J, Liu Q, Zhang X, et al. Magnetostratigraphy of a long Quaternary sediment core in the South Yellow Sea[J]. Quaternary Science Reviews, 2016, 144: 1-15. DOI:10.1016/j.quascirev.2016.05.025 |
| [45] |
田路萍, 李贵弘, 梁妮, 等. 环境中黑炭测定方法的研究进展[J]. 环境化学, 2020, 39(1): 166-178. Tian Luping, Li Guihong, Liang Ni, et al. Research progress of determination methods for environmental black carbon[J]. Environmental Chemistry, 2020, 39(1): 166-178. |
| [46] |
Lim B, Cachier H. Determination of black carbon by chemical oxidation and thermal treatment in recent marine and lake sediments and Cretaceous-Tertiary clays[J]. Chemical Geology, 1996, 131(1-4): 143-154. DOI:10.1016/0009-2541(96)00031-9 |
| [47] |
Wang X, Xiao J, Cui L, et al. Holocene changes in fire frequency in the Daihai Lake region(north-central China): Indications and implications for an important role of human activity[J]. Quaternary Science Reviews, 2013, 59: 18-29. DOI:10.1016/j.quascirev.2012.10.033 |
| [48] |
Wang X, Cui L, Xiao J, et al. Stable carbon isotope of black carbon in lake sediments as an indicator of terrestrial environmental changes: An evaluation on paleorecord from Daihai Lake, Inner Mongolia, China[J]. Chemical Geology, 2013, 347: 123-134. DOI:10.1016/j.chemgeo.2013.03.009 |
| [49] |
Wang X, Cui L, Yang S, et al. Stable carbon isotope records of black carbon on Chinese Loess Plateau since Last Glacial Maximum: An evaluation on their usefulness for paleorainfall and paleovegetation reconstruction[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2018, 509: 98-104. DOI:10.1016/j.palaeo.2017.08.008 |
| [50] |
Shao Y, Wyrwoll K-H, Chappell A, et al. Dust cycle: An emerging core theme in Earth system science[J]. Aeolian Research, 2011, 2(4): 181-204. |
| [51] |
Herring J R. Charcoal fluxes into sediments of the North Pacific Ocean: The Cenozoic record of burning[M]//Sundquist E T, Broecker W S. The Carbon Cycle and Atmospheric CO2: Natural Variations Archean to Present. New York: American Geophysical Union, 1985: 419-442.
|
| [52] |
崔敏, 陈颖军, 方引, 等. 北黄海泥质区多环芳烃与黑碳的沉积记录[J]. 海洋学报(中文版), 2015, 37(8): 39-46. Cui Min, Chen Yingjun, Fang Yin, et al. Sedimentary records of PAHs and BC from mud area of the central northern Yellow Sea China[J]. Acta Oceanologica Sinica, 2015, 37(8): 39-46. DOI:10.3969/j.issn.0253-4193.2015.08.004 |
| [53] |
姜晓华, 陈颖军, 唐建辉, 等. 渤海湾海岸带表层沉积物中黑碳的分布特征[J]. 生态环境学报, 2010, 19(7): 1617-1621. Jiang Xiaohua, Chen Yingjun, Tang Jianhui, et al. The distribution of black carbon in the surface sediments of coastal zone, Bohai Bay[J]. Ecology and Environmental Sciences, 2010, 19(7): 1617-1621. DOI:10.3969/j.issn.1674-5906.2010.07.018 |
| [54] |
Kang Y, Wang X, Dai M, et al. Black carbon and polycyclic aromatic hydrocarbons (PAHs) in surface sediments of China's marginal seas[J]. Journal of Oceanology and Limnology, 2009, 27(2): 297-308. DOI:10.1007/s00343-009-9151-x |
| [55] |
Xu X, Hong Y, Zhou Q, et al. Century-scale high-resolution black carbon records in sediment cores from the South Yellow Sea, China[J]. Journal of Oceanology and Limnology, 2018, 36(1): 114-127. DOI:10.1007/s00343-017-6214-2 |
| [56] |
方引, 陈颖军, 林田, 等. 黑碳在渤海泥质区的百年沉积记录[J]. 海洋学报(中文版), 2014, 36(5): 98-106. Fang Yin, Chen Yingjun, Lin Tian, et al. One hundred year sedimentary record of black carbon from mud area in Bohai Sea, China[J]. Acta Oceanologica Sinica, 2014, 36(5): 98-106. DOI:10.3969/j.issn.0253-4193.2014.05.011 |
| [57] |
Zhang J, Liu J, Wang H, et al. Characteristics and provenance implication of detrital minerals since Marine Isotope Stage 3 in Core SYS-0701 in the western South Huanghai Sea[J]. Acta Oceanologica Sinica, 2013, 32(4): 49-58. DOI:10.1007/s13131-013-0281-9 |
| [58] |
Liu J, Saito Y, Kong X, et al. Delta development and channel incision during marine isotope stages 3 and 2 in the western South Yellow Sea[J]. Marine Geology, 2010, 278(1-4): 54-76. DOI:10.1016/j.margeo.2010.09.003 |
| [59] |
Ai L, Han Z, Wu X, et al. Geochemical and grain-sized implications for provenance variations of the central Yellow Sea muddy area since the Middle Holocene[J]. Journal of Ocean University of China, 2020, 19(3): 577-588. DOI:10.1007/s11802-020-4211-0 |
| [60] |
Kong X, Zhang J, Bai W, et al. Provenance of sediments filling a paleo-channel that formed on the western Yellow Sea continental shelf during the last glacial period[J]. Journal of Ocean University of China, 2018, 17(6): 1309-1317. DOI:10.1007/s11802-018-3643-2 |
| [61] |
Yao Z, Shi X, Qiao S, et al. Persistent effects of the Yellow River on the Chinese marginal seas began at least~880 ka ago[J]. Scientific Reports, 2017, 7: 2827. DOI:10.1038/s41598-017-03140-x |
| [62] |
何梦颖, 梅西, 张训华, 等. 南黄海陆架区CSDP-1孔沉积物碎屑锆石U-Pb年龄物源判别[J]. 吉林大学学报(地球科学版), 2019, 49(1): 85-95. He Mengying, Mei Xi, Zhang Xunhua, et al. Provenance discrimination of detrital zircon U-Pb dating in the core CSDP-1 in the continental shelf of South Yellow Sea[J]. Journal of Jilin University(Earth Science Edition), 2019, 49(1): 85-95. |
| [63] |
Zhao Y, Yu Z, Chen F, et al. Vegetation response to Holocene climate change in monsoon-influenced region of China[J]. Earth-Science Reviews, 2009, 97(1-4): 242-256. DOI:10.1016/j.earscirev.2009.10.007 |
| [64] |
Miao Y, Zhang P, Lu S, et al. Late Quaternary pollen records from the Yangtze River Delta, East China, and its implications for the Asian monsoon evolution[J]. Arabian Journal of Geosciences, 2015, 8(10): 7845-7854. DOI:10.1007/s12517-015-1777-8 |
| [65] |
Zhang P, Miao Y, Zhang Z, et al. Late Cenozoic sporopollen records in the Yangtze River Delta, East China and implications for East Asian summer monsoon evolution[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2013, 388: 153-165. DOI:10.1016/j.palaeo.2013.08.014 |
| [66] |
Bird M I, Wynn J G, Saiz G, et al. The pyrogenic carbon cycle[J]. Annual Review of Earth and Planetary Sciences, 2015, 43: 273-298. DOI:10.1146/annurev-earth-060614-105038 |
| [67] |
Das O, Wang Y, Hsieh Y-P. Chemical and carbon isotopic characteristics of ash and smoke derived from burning of C3 and C4 grasses[J]. Organic Geochemistry, 2010, 41(3): 263-269. DOI:10.1016/j.orggeochem.2009.11.001 |
| [68] |
Schubert B A, Jahren A H. The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants[J]. Geochimica et Cosmochimica Acta, 2012, 96: 29-43. DOI:10.1016/j.gca.2012.08.003 |
| [69] |
Tipple B J, Meyers S R, Pagani M. Carbon isotope ratio of Cenozoic CO2: A comparative evaluation of available geochemical proxies[J]. Paleoceanography, 2010, 25(3): 1-11. |
| [70] |
Liu L, Song Y, Cui L, et al. Stable carbon isotopic composition of black carbon in surface soil as a proxy for reconstructing vegetation on the Chinese Loess Plateau[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2013, 388: 109-114. DOI:10.1016/j.palaeo.2013.08.012 |
| [71] |
Sage R F, Wedin D A, Li M. The biogeography of C4 photosynthesis: Patterns and controlling factors[M]//Sage R F, Monson R K. C4 Plant Biology. San Diego: Academic Press, 1999: 313-374.
|
| [72] |
Teeri J A, Stowe L G. Climatic patterns and the distribution of grasses in North America[J]. Oecologia, 1976, 23(1): 1-12. DOI:10.1007/BF00351210 |
| [73] |
Cavagnaro J B. Distribution of C3 and C4 grasses at different altitudes in a temperate arid region of Argentina[J]. Oecologia, 1988, 76(2): 273-277. DOI:10.1007/BF00379962 |
| [74] |
Hattersley P W. The distribution of C3 and C4 grasses in Australia in relation to climate[J]. Oecologia, 1983, 57(1): 113-128. |
| [75] |
殷立娟, 李美荣. 中国C4植物的地理分布与生态学研究Ⅰ. 中国C4植物及其与气候环境的关系[J]. 生态学报, 1997, 17(4): 16-29. Yin Lijuan, Li Meirong. A study on the geographic distribution and ecology of C4 plants in China Ⅰ. C4 plant distribution in China and their relation with regional climatic condition[J]. Acta Ecologica Sinica, 1997, 17(4): 16-29. |
| [76] |
王永吉, 吕厚远, 王国安, 等. C3, C4植物和现代土壤中硅酸体碳同位素分析[J]. 科学通报, 2000, 45(9): 978-982. Wang Yongji, Lü Houyuan, Wang Guo'an, et al. Carbon isotope analysis of silicates in C3, C4 plants and modern soil[J]. Chinese Science Bulletin, 2000, 45(9): 978-982. DOI:10.3321/j.issn:0023-074X.2000.09.016 |
| [77] |
饶志国, 贾国东, 朱照宇, 等. 中国东部表土总有机质碳同位素和长链正构烷烃碳同位素对比研究及其意义[J]. 科学通报, 2008, 53(17): 2077-2084. Rao Zhiguo, Jia Guodong, Zhu Zhaoyu, et al. Comparative study on carbon isotopes of total organic matter and long-chain n-alkanes in surface soils of Eastern China and their significance[J]. Chinese Science Bulletin, 2008, 53(17): 2077-2084. DOI:10.3321/j.issn:0023-074X.2008.17.013 |
| [78] |
Ode D, Tieszen L, Lerman J. The seasonal contribution of C3 and C4 plant species to primary production in a mixed prairie[J]. Ecology, 1980, 61(6): 1304-1311. DOI:10.2307/1939038 |
| [79] |
Yang S, Ding Z, Wang X, et al. Negative δ18O-δ13C relationship of pedogenic carbonate from Northern China indicates a strong response of C3/C4 biomass to the seasonality of Asian monsoon precipitation[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2012, 317-318: 32-40. DOI:10.1016/j.palaeo.2011.12.007 |
| [80] |
Boom A, Marchant R, Hooghiemstra H, et al. CO2- and temperature-controlled altitudinal shifts of C4- and C3-dominated grasslands allow reconstruction of palaeoatmospheric pCO2[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2002, 177(1-2): 151-168. DOI:10.1016/S0031-0182(01)00357-1 |
| [81] |
崔琳琳, 胡建芳, 王旭. 末次盛冰期以来中国南方C3/C4植被时空演化及影响机制[J]. 中国科学: 地球科学, 2019, 49(8): 1246-1258. Cui Linlin, Hu Jianfang, Wang Xu. Spatiotemporal evolution of C3/C4 vegetation and its controlling factors in Southern China since the Last Glacial Maximum[J]. Science China: Earth Sciences, 2019, 49(8): 1246-1258. |
| [82] |
Galy V, FranÇois L, France-Lanord C, et al. C4 plants decline in the Himalayan basin since the Last Glacial Maximum[J]. Quaternary Science Reviews, 2008, 27(13-14): 1396-1409. DOI:10.1016/j.quascirev.2008.04.005 |
| [83] |
Guo L. Linkage between C4 vegetation expansion and dune stabilization in the deserts of NE China during the Late Quaternary[J]. Quaternary International, 2019, 503: 10-23. DOI:10.1016/j.quaint.2018.10.026 |
| [84] |
Liu F, Chang X, Liao Z, et al. n-Alkanes as indicators of climate and vegetation variations since the last glacial period recorded in a sediment core from the northeastern South China Sea(SCS)[J]. Journal of Asian Earth Sciences, 2019, 171: 134-143. DOI:10.1016/j.jseaes.2018.09.018 |
| [85] |
Yang S, Ding Z, Li Y, et al. Warming-induced northwestward migration of the East Asian monsoon rain belt from the Last Glacial Maximum to the mid-Holocene[J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(43): 13178-13183. DOI:10.1073/pnas.1504688112 |
| [86] |
Yamada K, Ishiwatari R. Carbon isotopic compositions of long-chain n-alkanes in the Japan Sea sediments: Implications for paleoenvironmental changes over the past 85 kyr[J]. Organic Geochemistry, 1999, 30(5): 367-377. DOI:10.1016/S0146-6380(99)00012-1 |
| [87] |
Zhou B, Zheng H, Yang W, et al. Climate and vegetation variations since the LGM recorded by biomarkers from a sediment core in the northern South China Sea[J]. Journal of Quaternary Science, 2012, 27(9): 948-955. DOI:10.1002/jqs.2588 |
| [88] |
Wan S, Clift P D, Zhao D, et al. Enhanced silicate weathering of tropical shelf sediments exposed during glacial lowstands: A sink for atmospheric CO2[J]. Geochimica et Cosmochimica Acta, 2017, 200: 123-144. DOI:10.1016/j.gca.2016.12.010 |
| [89] |
万世明, 秦琳, 杨守业, 等. 南海冰期陆架风化与碳循环[J]. 第四纪研究, 2020, 40(6): 1532-1549. Wan Shiming, Qin Lin, Yang Shouye, et al. South China Sea shelf weathering in glacial periods and its link to carbon cycle[J]. Quaternary Sciences, 2020, 40(6): 1532-1549. |
| [90] |
Lu H, Liu W, Yang H, et al. 800-kyr land temperature variations modulated by vegetation changes on Chinese Loess Plateau[J]. Nature Communications, 2019, 10: 1958. DOI:10.1038/s41467-019-09978-1 |
| [91] |
Vidic N J, Montañez I P. Climatically driven glacial-interglacial variations in C3 and C4 plant proportions on the Chinese Loess Plateau[J]. Geology, 2004, 32(4): 337-340. DOI:10.1130/G20222.2 |
| [92] |
Zhang Z, Zhao M, Lu H, et al. Lower temperature as the main cause of C4 plant declines during the glacial periods on the Chinese Loess Plateau[J]. Earth and Planetary Science Letters, 2003, 214(3-4): 467-481. DOI:10.1016/S0012-821X(03)00387-X |
| [93] |
Uno K T, Polissar P J, Jackson K E, et al. Neogene biomarker record of vegetation change in Eastern Africa[J]. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(23): 6355-6363. DOI:10.1073/pnas.1521267113 |
| [94] |
Feakins S J, DeMenocal P B, Eglinton T I. Biomarker records of Late Neogene changes in Northeast African vegetation[J]. Geology, 2005, 33(12): 977-980. |
| [95] |
Fox D L, Koch P L. Tertiary history of C4 biomass in the Great Plains, USA[J]. Geology, 2003, 31(9): 809-812. |
| [96] |
An Z, Kutzbach J E, Prell W L, et al. Evolution of Asian monsoons and phased uplift of the Himalaya-Tibetan Plateau since Late Miocene times[J]. Nature, 2001, 411(6833): 62-66. DOI:10.1038/35075035 |
| [97] |
Lisiecki L E, Raymo M E. A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records[J]. Paleoceanography, 2005, 20(1): 1-17. |
| [98] |
Bond W J. Fires in the Cenozoic: A Late flowering of flammable ecosystems[J]. Frontiers in Plant Science, 2015, 5: 749. DOI:10.3389/fpls.2014.00749 |
| [99] |
Keeley J E, Rundel P W. Fire and the Miocene expansion of C4 grasslands: Miocene C4 grassland expansion[J]. Ecology Letters, 2005, 8(7): 683-690. |
| [100] |
Cai M, Fang X, Wu F, et al. Pliocene-Pleistocene stepwise drying of Central Asia: Evidence from paleomagnetism and sporopollen record of the deep borehole SG-3 in the western Qaidam Basin, NE Tibetan Plateau[J]. Global and Planetary Change, 2012, 94-95: 72-81. DOI:10.1016/j.gloplacha.2012.07.002 |
| [101] |
Sun Y, An Z. Late Pliocene-Pleistocene changes in mass accumulation rates of eolian deposits on the central Chinese Loess Plateau[J]. Journal of Geophysical Research, 2005, 110: D23101. DOI:10.1029/2005JD006064 |
| [102] |
Rea D K, Snoeckx H, Joseph L H. Late Cenozoic eolian deposition in the North Pacific: Asian drying, Tibetan uplift, and cooling of the Northern Hemisphere[J]. Paleoceanography, 1998, 13(3): 215-224. |
| [103] |
Sun Y, An Z, Clemens S C, et al. Seven million years of wind and precipitation variability on the Chinese Loess Plateau[J]. Earth and Planetary Science Letters, 2010, 297(3-4): 525-535. |
| [104] |
Seki O, Foster G L, Schmidt D N, et al. Alkenone and boron-based Pliocene pCO2 records[J]. Earth and Planetary Science Letters, 2010, 292(1-2): 201-211. |
| [105] |
顾家伟. 上新世以来苏北盆地与长江三角洲构造沉降史分析[J]. 地质科技情报, 2015, 34(1): 95-99+106. Gu Jiawei. Tectonic subsidence analysis of Subei Basin and Yangtze Delta from the Pliocene[J]. Geological Science and Technology Information, 2015, 34(1): 95-99+106. |
| [106] |
Li J, Fang X, Song C, et al. Late Miocene-Quaternary rapid stepwise uplift of the NE Tibetan Plateau and its effects on climatic and environmental changes[J]. Quaternary Research, 2014, 81(3): 400-423. |
| [107] |
张肖剑, 靳立亚, Andrey Ganopolski, 等. 3 Ma以来轨道尺度东亚冬季风演化的模拟研究[J]. 第四纪研究, 2020, 40(6): 1406-1417. Zhang Xiaojian, Jin Liya, Andrey Ganopolski, et al. Orbital variations of the East Asian winter monsoon during the past 3 Ma from a transient simulation[J]. Quaternary Sciences, 2020, 40(6): 1406-1417. |
| [108] |
鹿化煜, 张瀚之, 王逸超, 等. 渭河盆地新生代沉积序列与亚洲季风气候起源演化[J]. 第四纪研究, 2018, 38(5): 1057-1067. Lu Huayu, Zhang Hanzhi, Wang Yichao, et al. Cenozoic sedimentary sequence of Weihe Basin and origin and evolution of Asian monsoon climate[J]. Quaternary Sciences, 2018, 38(5): 1057-1067. |
| [109] |
程海, 李瀚瑛, 张旭, 等. 欧-亚-非大陆季风: 超级大陆与超级季风的雏形[J]. 第四纪研究, 2020, 40(6): 1381-1396. Cheng Hai, Li Hanying, Zhang Xu, et al. European-Asian-African continent: An early form of supercontinent and supermonsoon[J]. Quaternary Sciences, 2020, 40(6): 1381-1396. |
| [110] |
Ruddiman W F. Earth's Climate: Past and Future[M]. New York: W. H. Freeman and Company, 2001: 1-465.
|
| [111] |
郭正堂. 22-8 Ma风尘沉积记录的季风演变历史[M]//丁仲礼, 等编著. 中国西部环境演化集成研究. 北京: 气象出版社, 2010: 1-19. Guo Zhengtang. Monsoon evolution history recorded in the eolian deposits of 22-8 Ma[M]//Ding Zhongli, et al. eds. Study on Environmental Change of Western China. Beijing: China Meteorological Press, 2010: 1-19. |
| [112] |
Cerling T E, Wang Y, Quade J. Expansion of C4 ecosystems as an indicator of global ecological change in the Late Miocene[J]. Nature, 1993, 361(6410): 344-345. |
| [113] |
Pagani M. Late Miocene atmospheric CO2 concentrations and the expansion of C4 grasses[J]. Science, 1999, 285(5429): 876-879. |
| [114] |
Latorre C, Quade J, McIntosh W C. The expansion of C4 grasses and global change in the Late Miocene: Stable isotope evidence from the Americas[J]. Earth and Planetary Science Letters, 1997, 146(1-2): 83-96. |
| [115] |
Ecker M, Kelley D, Sato H. Modelling the effects of CO2 on C3 and C4 grass competition during the mid-Pleistocene transition in South Africa[J]. Scientific Reports, 2020, 10(1): 16234. DOI:10.1038/s41598-020-72614-2 |
| [116] |
王萍, 殷立娟, 李建东. 东北草原区C3、C4植物的生态分布及其适应盐碱环境的生理特性[J]. 应用生态学报, 1997, 8(4): 407-411. Wang Ping, Yin Lijuan, Li Jiandong. Ecological distribution and physiological characteristics of C3 and C4 plants adapted to saline-alkali environment in the grasslands of Northeast China[J]. Chinese Journal of Applied Ecology, 1997, 8(4): 407-411. |
| [117] |
殷立娟, 王萍. 中国东北草原植物中的C3和C4光合作用途径[J]. 生态学报, 1997, 17(2): 3-13. Yin Lijuan, Wang Ping. Distribution of C3 and C4 photosynthetic pathways of plants on the steppe of Northeastern China[J]. Acta Ecologica Sinica, 1997, 17(2): 3-13. |
| [118] |
Wang P, Tian J, Cheng X. Transition of Quaternary glacial cyclicity in deep-sea records at Nansha, the South China Sea[J]. Science in China (Series D: Earth Sciences), 2001, 44(10): 926-933. |
| [119] |
Li L, Li Q, Tian J, et al. A 4-Ma record of thermal evolution in the tropical western Pacific and its implications on climate change[J]. Earth and Planetary Science Letters, 2011, 309(1-2): 10-20. |
| [120] |
Herbert T D, Lawrence K T, Tzanova A, et al. Late Miocene global cooling and the rise of modern ecosystems[J]. Nature Geoscience, 2016, 9(11): 843-849. |
| [121] |
Wu B, Wu N. Terrestrial mollusc records from Xifeng and Luochuan L 9 loess strata and their implications for paleoclimatic evolution in the Chinese Loess Plateau during Marine Oxygen Isotope Stages 24-22[J]. Climate of the Past, 2011, 7(2): 349-359. |
| [122] |
Guo Z, Biscaye P, Wei L, et al. Summer monsoon variations over the last 1.2 Ma from the weathering of loess-soil sequences in China[J]. Geophysical Research Letters, 2000, 27(12): 1751-1754. |
| [123] |
Wang H, Lu H, Zhao L, et al. Asian monsoon rainfall variation during the Pliocene forced by global temperature change[J]. Nature Communications, 2019, 10: 5272. DOI:10.1038/s41467-019-13338-4 |
2 Laboratory for Marine Geology, Pilot National Laboratory for Marine Science and Technology(Qingdao), Qingdao 266237, Shandong;
3 University of Chinese Academy of Sciences, Beijing 100049;
4 China Offshore Fugro Geosolutions(Shenzhen) Co., Ltd., Shenzhen 518068, Guangdong)
Abstract
Terrestrial vegetation is an important part of the global ecosystem. The C3/C4 evolution has great implication for our understanding the response and adaptation mechanisms of ecosystem to global changes. The timing, processes and mechanism of C4 plant expansion in East Asia since the Late Pliocene are still controversial. Especially, the evolution history of C3/C4 plant in the Yangtze River Basin at the middle latitude and its comparison with plant evolution through different latitude regions of East Asia remains unclear. Here we firstly reported the black carbon record from the Continental Shelf Drilling Program(CSDP) Core CSDP-1(34°18'N, 122°22'E; water depth 52.5m; 300.10m in length and since 3.5Ma) from the South Yellow Sea drilled by Research Vessel KAN 407 in June 2013, as well as surface sediment samples of seven major rivers in Eastern China. The content, mass accumulation rate(MAR) and carbon isotopic composition of black carbon extracted from sediments of core CSDP-1, were used to reconstruct the C3/C4 vegetation evolution in Eastern China since the Late Pliocene. During the Late Pliocene(ca. 3.0~2.6Ma), the carbon isotopic composition of black carbon of the study core shows more positive values, indicating the increased relative abundance of C4 plants in the Yangtze River Basin during that period. We suggest that such evolution was possibly controlled by the increased aridity in East Asia and the decline of atmospheric CO2 concentration at the Late Pliocene. In contrast, reduced C4 vegetation abundance in the mid-high latitude regions(i.e., the Loess Plateau) in the Late Pliocene likely suggests that lower temperature became the limited factor for growth of C4 vegetation. Since the Middle Pleistocene(ca.1.0Ma), the carbon isotope composition of black carbon at core CSDP-1 reversed to more negative, suggesting that the relative abundance of C4 vegetation in the source area declined at that period. Although the sediment source of the study area shifted from the Yangtze River to the Yellow River located at higher latitude since about 0.8Ma, similar negative trend of carbon isotope composition from different latitudes of East Asia suggests that it's a regional phenomenon of C4 vegetation evolution, which was likely caused by global cooling since the Middle Pleistocene. A comprehensive review suggests the very different evolution trend of C3/C4 plant in different latitude regions of East Asia from modern to million year time-scale. Specifically, stronger aridity of the low-middle latitude regions would preferably increase the abundance of C4 plant, while low temperature in middle-high latitude regions is more critical in limiting the growth of C4 plant. Therefore, this study highlights the significant influence of regional climate(i.e., temperature and aridity) in C3/C4 vegetation evolution in East Asia, although we cannot exclude the possible role of atmospheric CO2 level.