 2016, Vol.36
2016, Vol.36



②. 中国科学院大学, 北京 100049;
③. 北京师范大学地理学与遥感科学学院, 北京 100875)
历史气候定量重建是过去全球变化研究计划(PAGES)的一项重要内容,已成为全球变化研究中的前沿课题之一。IPCC第五次评估报告(AR5)也考虑将历史气候研究成果作为气候变化研究的新证据[1]。物候学作为研究周期性生物事件发生时间、 影响因子以及同种或不同种间物候期相互关系的科学[2]在探讨生态系统对气候变化响应上起了重要作用。由于自然物候期变化主要受环境因素特别是气候变化的影响,同时相比于沉积、 冰芯等代用资料,物候资料具有客观性强、 分辨率高等特点,因此物候资料在重建历史气候变化中具有重要作用[1]。早在20世纪70年代,竺可桢[3]便主要利用物候学资料和方法重建了中国过去5000年的气候变化。这之后,许多学者根据历史文献中的物候记录,重建了不同区域、 时长和分辨率的历史气候序列[4~6]。在物候学研究中,物候模型是一种重要的参数化和定量化研究手段。尤其是在历史时期,缺乏实际观测资料的情况下,物候模型是唯一能够重建过去和预测未来物候变化的方法[7]。通过物候模型可以基于植物的生理机制建立环境因子与植物物候期之间的数学关系,进而根据不同时期植物物候的特征反演气候的变化阶段。随着物候模型研究的深入,众多学者通过比较多种模型的模拟效果,评估了各模型针对不同区域和不同物种的适用性,为区域性的气候定量重建奠定了基础[8~10]。
现有物候模型以植物物候模型居多。由于北半球中高纬度地区木本植物物候主要受气温影响[11~13],在此基础上综合考虑光照、 水分等因素可较为准确的模拟木本植物物候期。截止目前,木本植物物候模型的发展较为成熟,在气候定量重建中的应用也十分广泛。例如Aono和Saito[14]基于224年的日本樱花始花期数据,利用物候模型重建了日本京都地区9~14世纪3月份平均温度,识别了包含“中世纪暖期”在内的多个气候变化阶段。Chuine等[6]运用基于过程的物候模型,将法国Burgundy地区葡萄收获日期转化为温度异常值,重建了1370~2003年间法国东部春夏季气温异常值。该重建序列表明2003年气候的异常增暖为前所未有的。相比于木本植物,草本植物与气候要素的关系较为复杂,温度对物候的作用并不明显; 不同区域草本植物受到地理条件和小气候的影响,对气候要素的响应规律也存在较大差异[15]。总体而言,草本植物模型难以较为准确的模拟植物与气候变化的相关关系,利用草本模型定量重建历史气候的研究也未见报道。
本文系统综述国内外现有主要木本植物物候模型,评述各模型的主要特点,详细介绍植物物候模型在气候定量重建中的应用,再总结并展望了未来研究中需要注意的关键问题和发展方向,希冀为利用物候资料进行全球变化研究提供方法依据和参考。
2 物候模型物候模型建立的基础是植物的遗传特性、 生长发育规律及其对环境因子的响应关系。这3个因素共同决定了植物的生长过程和物候期的发生和持续时间[16, 17]。在北半球中高纬度地区,植物春季物候的提前和秋季的推迟趋势都主要受温度变化影响[18~21]。因此,物候模型常被用于重建历史温度的变化。现有应用于气候定量重建中的模型可分为两类: 统计模型和过程模型。
2.1 统计模型统计模型也称回归模型,主要是利用物候期与气象因子之间的统计关系建立回归方程。这种模型不考虑植物生理生态过程,是研究物候与气候关系直接有效的方法[22]。统计模型有许多类型,其中一种类型利用物候期与每年不同时期气温的相关性建立它们间的线性回归方程[23]。这种方法在重建中的应用不多,因为植物物候对温度的响应往往呈非线性的关系,简单线性回归方程无法准确描述这种关系。另一种类型的统计模型则较为复杂并且常见于气候定量重建研究中。典型例子是Schwartz等[24, 25]提出的预测洲际尺度春季物候期的多元回归模型:
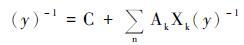
|
(1) |
公式(1)中,y是物候事件发生的日期; C和AK是常数,XK是特定的自变量,通常包括从1月1日起的积温(以-0.6度为阈值)、 某一温度范围内时间的累积值以及特定天气的日数来衡量植被对温度的非线性响应。考虑到植物需要达到一定的冷激需求才能够发芽,之前的模型默认了在1月1日时冷激需求已经被满足。Schwartz等[24, 25]又加入了低温累积和(小于7.2℃的温度和)作为参数。该模型由于考虑了较多参数,并有生理学方面的考虑,因此作为春季物候期的代表,被应用于美国乃至北半球[26]。
2.2 过程模型过程模型是一种半机理性模型,其建立基础是生物过程与环境因子之间的因果关系,是指通过植物每日发育进度与环境因子的关系,利用数学模型模拟植物的生长过程,这种因果关系必须是经过充分检验[22]。根据物候模型模拟的植物的不同生理阶段,可将过程模型分为单过程模型和多过程模型。
2.2.1 单过程模型单过程模型考虑春季植物打破生态休眠期所需生长驱动单元的累积量[27]。在历史气候定量重建中主要应用的单过程模型有两种,第一种单过程模型被称为热时(thermal time)模型[28]或春季积温(spring warming,简称SW)模型[29]。该模型的理论基础是Reaumur[30]提出的积温理论,即植物的发育进度和物候期是由过去一段时间的累积温度所决定的。SW模型是最简单的植被物候过程模型,只有3个参数: t0、 Tb1和F*。SW的公式如以下(2)和(3)所示:

|
(2) |
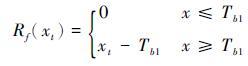
|
(3) |
其中,y是物候事件发生的时间,xt是第t天的日平均气温,Rf(xt)是驱动单元; Tb1是临界温度,t0是驱动单元开始累积的时间; F*指发育状态的阈值,当累积温度达到这个阈值时物候事件就发生。
另一种模型是由Aono和Omoto[31]建立的温度变换日数(the number of days transformed to standard temperature,简称DTS)模型。它借鉴化学反应速率随温度变化关系的经验公式阿伦尼乌斯公式(Arrhenius equation)来定义物候驱动单元的函数形式[31]。模型假设植物每一天的生长速率受当日温度影响,每日的DTS值((ts)ij)可以用该日生长速率和标准温度下的生长速率比值来表示,如公式(4)所示:
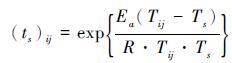
|
(4) |
其中,Tij表示第i年第j天的日平均温度,Ts表示标准温度,常取288.2K,Ea(J/mol)是温度特征值,R是普适气体常量(值为8.314J/(mol·K))。
当累积的DTS值达到多年平均值DTSN时,Bi即是物候期发生的时间(如公式5),其中N是用于平均的年份,常取25或30[5, 31]; D表示积温的开始日期。
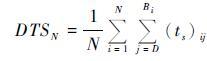
|
(5) |
随着植物学和物候学研究的深入,Taiz和Zeiger[17]在实验中发现,休眠期是木本植物适应寒冷气候,保证生长发育的一个重要阶段。许多木本植物的发育过程都需要经过休眠期的冷刺激积累,冷刺激进度的不同也会导致休眠期打破时间的差异。这一观点在后来的研究中也逐渐发展成为最常见的多过程模型[32, 33]。总体上来说,这类模型对中高纬度地区的多数木本植物物候模拟的整体效果好于简单的统计模型和SW模型[34],但目前在气候重建中还没有相应的应用。迄今为止,国际上气候定量重建中应用最多的多过程模型是欧洲的葡萄收获日期模型。
葡萄收获日期模型主要用于模拟葡萄成熟期的物候变化过程[6],共包含3个阶段,即开花阶段采用SW模型模拟开花期、 开花后进入到转熟阶段(葡萄果实变色过程)以及该阶段又可以分为细胞伸长阶段C*和光合产物积累阶段P*。后两个阶段采用类似积温的算法(如公式6和7),但驱动单元分别为三次多项式和指数函数(如公式8和9)。

|
(6) |
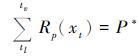
|
(7) |
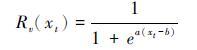
|
(8) |
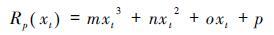
|
(9) |
其中,xt表示第t日的平均温度,tf和tl分别是细胞成长阶段C*的开始和结束时间,tl同时也是光合产物积累阶段P*的开始时间,tv是P*的结束时间; Rv(xt)和Rp(xt)分别是两个阶段的驱动单元; a、 b、 m、 n、 o和p是模型的拟合参数。
3 物候模型在气候定量重建中的应用利用物候模型定量重建气候的基础是植物对气候变化的响应关系。将气候数据输入到物候模型中可以输出物候期,反过来也可以通过观测到的物候期推测气候变化。利用物候模型重建历史气候变化的步骤主要包括以下3步: 1)通过模型对比,选择模拟效果最好的物候模型; 2)根据现代观测的物候数据和平均温度等气候数据确定物候模型的参数; 3)将历史物候数据带入物候模型,得到重建的气候数据。
上文中介绍的几种物候模型在国内外的历史气候定量重建上都有许多成功案例。
3.1 统计模型在温度重建中的应用统计模型作为最简单的物候模型,在国际上被广泛用来定量重建温度变化。Možný等[35]利用1845年以来系统观测的谷物收获日期以及从编年史、 日记、 财政账目中提取的谷物收获日期,建立了1501~2008年小麦(wheat)、 黑麦(rye)、 大麦(barley)、 燕麦(oats)4种谷物的物候序列,经计算上述4种作物的收获日期与冬小麦的收获日期都有较高的相关性,因此将这4种物候期转化为冬小麦的物候期用以重建捷克1501~2008年3~6月的温度。从重建的温度序列中可准确识别1个寒冷时段(1659~1705年)和3个温暖时段(1517~1542年、 1788~1834年和1946~2008年)。从重建精度上来看,重建的序列可以解释70%的温度变化,误差为0.50~0.56℃[35](图 1)。

|
图 1 基于冬小麦收获期重建的1501~2008年捷克地区3~6月平均温度距平(蓝线)及其30年的滑动平均值(黑线)引自文献[35],灰色区域表示95%的置信区间,红色线条表示在1848~2006年测量的温度距平的30年滑动平均值 Fig. 1 Reconstructed mean Czech March-June temperature anomalies(blue curve)based on winter wheat harvest days in the period 1501~2008 and smoothed by Gaussian filter for 30 years(black),cited from reference[35]. Gray area approximates to 95% confidence interval and red curve indicates the measured temperature anomalies smoothed by Gaussian filter for 30 years in 1848~2006 |
统计模型也是国内的气候定量重建中用到的最主要的物候模型。例如,郑景云等[36]根据华中地区的历史物候记录,重建了1850~2008年华中地区年均气温的变化序列。值得一提的是,与一般的统计模型不同,该研究除了利用物候记录外,还结合了清代档案中的降雪日数记录和5个树轮宽度年表,采用逐步回归方法,建立了温度和各代用资料的多元回归模型,重建的序列可以解释66%的温度变化[36]。这一研究为物候模型定量重建气候变化提供了新的思路,即结合物候和其他多种代用资料共同指示温度变化过程。
3.2 SW模型在芬兰地区温度重建中的应用SW模型主要在芬兰地区被用于验证历史气象观测数据。Linkasalo等[37]基于芬兰地区1846~2005年8种木本植物的开花期和芽开放期的物候观测记录,重建了8条物候序列。再结合SW模型重建了长达160年的春季温度变化序列,最后利用重建的温度序列对气象观测数据进行验证。重建的温度序列表明芬兰的春季温度平均每个世纪增长1.8℃,这与气象观测的1.5℃/10a非常接近(图 2)。SW模型的模拟效果明显要好于统计模型,这表明基于植物生长机理的模型能更好地揭示气候因素对植物物候的影响机制[10]。但是对于不同的物种而言,重建的升温速率有明显差别,这说明利用SW模型重建历史温度的精确度受所选用于重建的物种类型影响,例如,利用欧洲花楸(Sorbus aucuparia,1.4℃/10a)重建的芬兰地区春季升温速率显然比辽东桤木(Alnus incana,2.4℃/10a)更准确(图 2)。对于同一物种来说,不同物候期也决定了重建的效果。从图 2中可以看出利用欧洲山杨(Populus tremula)芽开放期重建的升温速率(1.75℃/10a)比基于开花期重建的升温速率(0.7℃/10a)要更接近与观测数据[37]。

|
图 2 利用温度计观测方法和物候记录重建方法获得的升温速率(℃/10a)(1846~2005年)引自文献[37],虚线和灰色阴影面积分别表示观测到的升温速率平均值和95%的置信区间。柱状图和短竖线分别表示利用芽开放期(灰色)和开花期(白色)重建的升温速率及其95%的置信区间 Fig. 2 Temperature increases(℃/10a)according to thermometer measurements during spring and estimates based on phenological records(1846~2005),cited from reference[37]. Where dashed line and grey shadow area represent the mean temperature increases and 95% confidence intervals,respectively. Bars and vertical lines show the estimated temperature increases by leaf bud burst(grey)and flowering dates(white)and their 95% confidence intervals,respectively |
3.3 DTS模型在日本地区温度重建中的应用
从1996年开始,DTS模型就被日本气象局广泛的应用到樱花花期的预报中[31, 38]。研究发现,在预报20世纪的樱花盛花期上,DTS模型的误差比SW模型小0.4天[5]。因此,DTS模型也常被用于定量重建日本地区的气候变化。例如,Aono和Kazui[5]利用日本历史文献,在前人研究基础上整理形成了京都地区732年的樱花盛花期序列。首先根据现代的樱花盛花期观测数据和温度数据,确定了积温开始日期D和温度特征值Ea(见公式4和5)。然后将多年平均气温序列作为基准(1961~1990年),在基准气温序列上增加一个ΔT, 使得模拟的樱花盛花期与观测值一致(如公式10)。基准气温加上ΔT即为重建的气温,公式(10)如下所示:

|
(10) |
重建的结果识别了9世纪以来京都地区的4个寒冷时段。同时重建温度的年代际变化与太阳黑子的活动周期相吻合,揭示了太阳活动对温度变化的影响机制[5]。
利用类似的方法,Aono和Saito[14]从日记、 编年史、 诗歌中提取出日本樱花开花时间,并收集紫藤(Wisteria floribunda)、 日本杏(Prunus mume)、 日本海棠(Kerria japonica)、 木牡丹(Paeonia suffruticosa)等物候期合并成为一个224年的物候序列,结合DTS模型重建了京都地区9~14世纪的3月份平均气温。结果表明京都地区10世纪的温度比现在高,此后温度有所下降,到1180s温度开始回暖,但在1310s后的整个14世纪中期温度再次降低[14]。
总体上来看,DTS模型在日本地区对樱花花期的模拟效果良好,在温度重建上的应用已经比较成熟,重建温度的均方根误差可达到小于0.1℃[5](图 3),是目前国内外重建精度最高的。

|
图 3 DTS模型重建的3月均温与实际温度的31年滑动平均值(引自文献[5]) Fig. 3 Estimated March mean temperature and actual temperature during the instrumental period,smoothed using a 31-year-time span,cited from reference[5] |
3.4 葡萄收获日期模型在欧洲温度重建中的应用
欧洲地区的葡萄收获日期的记录十分丰富,目前发现的最早的有连续记录的葡萄收获日期可追溯到1370年[39]。葡萄成熟与温度变化的关系也十分紧密,因此,葡萄的收获期常被用作重建欧洲地区的温度变化。
例如,Chuine等[6]利用葡萄的多过程模型和法国勃艮第(Burgundy)地区的葡萄(Pinot Noir)收获日期的历史观测记录,重建了1370~2003年春夏的温度变化。重建结果表明勃艮第地区自1370年,气温曾经多次达到20世纪90年代的高度。但是2003年比较异常,很可能是自1370年以来气温的最高值(图 4)。将该重建结果和树轮等其他方法重建的温度序列作线性相关分析,相关系数可达到0.45~0.75[6]。

|
图 4 法国勃艮第地区基于葡萄收获日期重建的1370~2003年4~8月温度距平(引自文献[6])黑色曲线表示重建的温度距平,黄色曲线表示距平30年的滑动平均值; 紫色曲线表示置信区间,蓝色阴影面积表示置信区间的11年的滑动平均值;橙色曲线表示观测的站点数量;红线表示重建温度2个标准差的范围(1901~2003年);红色和蓝色的箭头分别指示温暖和寒冷的时期 Fig. 4 April-August temperature anomalies in Burgundy,France,as reconstructed from grape harvest dates from 1370 to 2003,cited from reference[6]. Where black curve and yellow curve represent reconstructed anomalies and the 30-year Gaussian filter,respectively. Purple curve and blue shaded area are the confidence interval and the 11-year filter. Orange curve shows the numbers of the stations. Red lines exhibit the 2σ interval of the reconstructed temperature(1901~2003). Red and blue arrows indicate the warm and cold periods,respectively |
在瑞士地区,也有利用葡萄收获日期重建温度变化的案例。Meier等[40]利用回归模型和瑞士地区1480~2006年的葡萄收获日期记录,重建了该地区近500年的4~8月温度距平。结果发现瑞士地区17世纪和19世纪初期较为寒冷,而在18世纪初期和近代较为温暖。
但是,利用葡萄重建温度变化仍存在很多不确定性[41]。首先,从上述两个重建工作可以看出,虽然是利用同一物种对温度进行重建,但所选取的模型并不相同。这主要是由于不同地区葡萄收获期对气候变化的响应机制存在差异。从重建效果上来看,Meier等[40]的重建误差达到0.82℃,与日本基于樱花的温度重建相比较高[5]。这表明现有模型还不能十分准确描述葡萄收获期和温度变化的相关关系。一方面是葡萄栽培技术变化和其他的非气候因素对重建工作产生了一定的影响,例如人工的施肥、 修剪和耕作等都会改变葡萄的自然生长状况,而这种影响也很难被模型模拟出来; 另一方面,不同的葡萄物种对气候变化的响应敏感性存在差异。如果在模型定标和模拟重建中所用的葡萄品种不同,也会进一步影响重建的准确性。
4 结论和讨论从应用情况上来看,多种物候模型在国际上已被广泛的应用到气候的定量重建中,但在国内重建研究中所利用的物候模型还局限于统计模型,这体现了国内外研究的差距。同时,现有物候模型仅被用于重建温度序列,还未有研究利用物候模型重建其他气候因子的变化序列。然而,随着植物物候研究不断深入,光照和降水对物候期的影响作用已经在很多地区被证实[42, 43]。在亚热带地区,温暖的冬季已经很难满足植物发育的温度要求,光周期代替温度对诱导植物开花起主要作用[44]。在干旱半干旱区域,水分条件是植物生长的主要限制因子[12],因此,降水对植物物候的影响要更加明显[45, 46]。在这些地区,部分学者尝试在物候模型中加入光照和降水的参数。例如,Schaber和Badeck[47]首次将光周期加入到过程模型中,建立了促进-抑制模型; Blümel和Chmielewski[48]在SW模型基础上进行了改进,增加了白天时长对植物生长的影响,该模型认为白天时长越长,植物对温度的利用效率越高; Chen等[49]在对青藏高原地区的植物物候进行模型模拟时,假设降水对植物春季物候的诱导机制和温度相类似,植物物候期与过去一段时间内的累积降水量相关。仿照SW模型建立了降水和温度共同影响的多过程模型。以上这些模型应用的结果在一定程度上考虑了多种气候因素对物候的综合影响,但是由于目前在影响机理上仍然尚不明确,模型的模拟精度并不高。这些模型在气候定量重建中有十分重要的潜在价值,为今后建立更准确的物候模型,重建历史降水等气候因子变化序列奠定了基础。
总体而言,物候模型研究在不断完善中。在未来的研究中还需要在以下几个方面重点关注和加强:
(1) 利用物候模型定量重建气候的关键问题是模型的选择。对于不同物种和地区,模型的适用性具有明显的差异。随着对植物生长和环境要素影响机理的深入了解,植物物候模型越来越复杂,但并不意味着越复杂的模型对所有物种的模拟效果越好。因此,重建工作不能仅仅套用前人研究中的模型,而是应该对多模型进行比较,选择最优模型。
(2) 采用物候模型定量重建历史气候的各种方法均以气候要素、 特别是温度要素驱动某物候期为前提条件。然而,物候期对气候变化的响应关系可能处于变动之中。最新研究表明[50],物候期对气温的敏感性就有一定波动,这对重建结果或有重要影响。因此,在后续采用物候模型定量重建气候的工作中,尚需深入考虑该因素,评价其影响程度和所引起的不确定性。
(3) 要详细鉴定历史物候记录中的物种是否与现代观测的物种一致。物种的鉴别可以依据植物的地理分布和物候规律进行推断[51]。为了便于记录和分析,同一属的不同种植物可分开进行种植和观测,以防混淆[41]。
(4) 本文所介绍的模型主要用于春季和夏季物候期,诸如展叶、 开花和果实成熟期的预测,却很少涉及秋季物候模型。因此,重建的气候序列也以春季和夏季为主。目前大多数秋季物候模型诸如预测叶衰老的模型,均基于一些经验关系[52, 53],或者气温的临界点(假设叶片衰老在达到某个极端低温阈值时触发)。也有模型通过叶片碳平衡假设预测叶衰老[54]。Delpierre等[55]尝试性的提出了基于气温和光周期共同控制的过程模型预测叶变色期。但这些方法预测的准确性缺乏广泛彻底的验证。这一方面是由于秋季物候比春季物候更加易变[56],缺乏对秋季物候生理机制方面的研究; 另一方面,秋季物候相对于春季物候难以观测,例如,从观测者的视角很难准确的把握叶变色的程度、 落叶期很容易受到风的干扰,等等。观测数据以及观测的准确性影响了秋季物候模型的发展。为建立准确的秋季物候模型,需要继续加强秋季物候的观测以及秋季物候生理机制的研究。这对更清晰理解秋季物候变化过程,对更充分利用大量秋季资料进行历史气候重建也显得非常关键。
| 1 |
IP CC.
Summary for Policymakers. Climate Change 2013:The Physical Science Basis. In:Stocker T F, Qin D H, Plattner G et al. eds. Contribution of Working Group Ⅰ to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change[M]. Cambridge: Cambridge University Press, 2013 : 3 ~29.
( 0) 0)
|
| 2 |
Lieth H.
Phenology and Seasonality Modeling[M]. Berlin: Springer Verlag, 1974 : 1 ~444.
(0)
|
| 3 |
竺可桢, 宛敏渭.
物候学[M]. 北京: 科学出版社, 1973 : 1 ~173.
Zhu Kezhen, Wan Minwei. Phenology[M]. Beijing: Science Press, 1973 : 1 ~173. ( 0)
|
| 4 |
Ge Q S, Zheng J Y, Fang X Q, et al. Winter half-year temperature reconstruction for the middle and lower reaches of the Yellow River and Yangtze River, China, during the past 2000 years[J].
The Holocene,2003, 13 (6) : 933~940.
(0)
|
| 5 |
Aono Y, Kazui K. Phenological data series of cherry tree flowering in Kyoto, Japan, and its application to reconstruction of spring time temperatures since the 9th century[J].
International Journal of Climatology,2008, 28 (7) : 905~914.
(0)
|
| 6 |
Chuine I, Yiou P, Viovy N, et al. Historical phenology:Grape ripening as a past climate indicator[J].
Nature,2004, 432 (7015) : 289~290.
(0)
|
| 7 |
Cleland E E, Chuine I, Menzel A, et al. Shifting plant phenology in response to global change[J].
Trends in Ecology & Evolution,2007, 22 (7) : 357~365.
(0)
|
| 8 |
Kramer K, Leinonen I, Loustau D. The importance of phenology for the evaluation of impact of climate change on growth of boreal, temperate and Mediterranean forests ecosystems:An overview[J].
International Journal of Biometeorology,2000, 44 (2) : 67~75.
(0)
|
| 9 |
Matzneller P, Blümel K, Chmielewski F M. Models for the beginning of sour cherry blossom[J].
International Journal of Biometeorology,2014, 58 (5) : 703~715.
(0)
|
| 10 |
张爱英, 王焕炯, 戴君虎, 等. 物候模型在北京观赏植物开花期预测中的适用性[J].
应用气象学报,2014, 25 (4) : 483~492.
Zhang Aiying, Wang Huanjiong, Dai Junhu, et al. Applicability analysis of phenological models in flowering time prediction of ornamental plants in Beijing area[J]. Journal of Applied Meteorological Science,2014, 25 (4) : 483~492. ( 0)
|
| 11 |
Schwartz M D.
Phenology:An Integrative Environmental Science[M]. Dordrecht: Springer, 2003 : 1 ~610.
(0)
|
| 12 |
阿布都克日木·阿巴司, 王荣梅, 阿不都西库尔·阿不都克力木. 1982-2010年喀什木本植物物候变化与气候变化的关系[J].
第四纪研究,2013, 33 (5) : 927~935.
Abudoukerimu·Abasi, Wang Rongmei, Abudouxiku'er·Abudoukelimu. The relationship with woody plants phonological variation characters and climatic change from 1982 to 2010 in Kashi[J]. Quaternary Sciences,2013, 33 (5) : 927~935. ( 0)
|
| 13 |
方修琦, 萧凌波, 葛全胜, 等. 湖南长沙、衡阳地区1888-1916年的春季植物物候与气候变化[J].
第四纪研究,2005, 25 (1) : 74~79.
Fang Xiuqi, Xiao Lingbo, Ge Quansheng, et al. Changes of plants phenophases and temperature in spring during 1888-1916 around Changsha and Hengyang in Hunan Province[J]. Quaternary Sciences,2005, 25 (1) : 74~79. ( 0)
|
| 14 |
Aono Y, Saito S. Clarifying springtime temperature reconstructions of the medieval period by gap-filling the cherry blossom phenological data series at Kyoto, Japan[J].
International Journal of Biometeorology,2010, 54 (2) : 211~219.
(0)
|
| 15 |
符瑜, 潘学标. 草本植物物候及其物候模拟模型的研究进展[J].
中国农业气象,2011, 32 (3) : 319~325.
Fu Yu, Pan Xuebiao. Herb phenology and progress of phenological models[J]. Chinese Journal of Agrometeorology,2011, 32 (3) : 319~325. ( 0)
|
| 16 |
Böhlenius H, Huang T, Charbonnel-Campaa L, et al. CO/FT regulatory module controls timing of flowering and seasonal growth cessation in trees[J].
Science,2006, 312 (5776) : 1040~1043.
(0)
|
| 17 |
Taiz L, Zeiger E.
Plant Physiology[M]. Sunderland: Sinauer Associates, 2006 : 1 ~700.
(0)
|
| 18 |
Wolfe D W, Schwartz M D, Lakso A N, et al. Climate change and shifts in spring phenology of three horticultural woody perennials in Northeastern USA[J].
International Journal of Biometeorology,2005, 49 (5) : 303~309.
(0)
|
| 19 |
Menzel A, Sparks T H, Estrella N, et al. European phenological response to climate change matches the warming pattern[J].
Global Change Biology,2006, 12 (10) : 1969~1976.
(0)
|
| 20 |
Ge Q S, Wang H J, Rutishauser T, et al. Phenological response to climate change in China:A meta-analysis[J].
Global Change Biology,2015, 21 (1) : 265~274.
(0)
|
| 21 |
Dai J H, Wang H J, Ge Q S. Multiple phenological responses to climate change among 42 plant species in Xi'an, China[J].
International Journal of Biometeorology,2013, 57 (5) : 749~758.
(0)
|
| 22 |
Chuine I, Kramer K, HNninen H.
Plant development models. In:Schwartz M D ed. Phenology:An Integrative Environmental Science[M]. Dordrecht: Springer, 2003 : 217 ~235.
(0)
|
| 23 |
Emberlin J, Detandt M, Gehrig R, et al. Responses in the start of Betula(birch)pollen seasons to recent changes in spring temperatures across Europe[J].
International Journal of Biometeorology,2002, 46 (4) : 159~170.
(0)
|
| 24 |
Schwartz M D, Reiter B E. Changes in North American spring[J].
International Journal of Climatology,2000, 20 (8) : 929~932.
(0)
|
| 25 |
Schwartz M D, Marotz G A. An approach to examining regional atmosphere plant interactions with phenological data[J].
Journal of Biogeography,1986, 13 (6) : 551~560.
(0)
|
| 26 |
Schwartz M D, Ahas R, Aasa A. Onset of spring starting earlier across the Northern Hemisphere[J].
Global Change Biology,2006, 12 (2) : 343~351.
(0)
|
| 27 |
Wang Huanjiong, Dai Junhu, Ge Quansheng. The spatiotemporal characteristics of spring phenophase changes of Fraxinus chinensis in China from 1952 to 2007[J].
Science China:Earth Sciences,2012, 55 (6) : 991~1000.
(0)
|
| 28 |
Cannell M G R, Smith R I. Thermal time, chill days and prediction of budburst in Picea sitchensis[J].
Journal of Applied Ecology,1983, 20 (3) : 951~963.
(0)
|
| 29 |
Hunter A F, Lechowicz M J. Predicting the timing of budburst in temperate trees[J].
Journal of Applied Ecology,1992, 29 (3) : 597~604.
(0)
|
| 30 |
Reaumur R A.
Observations du thermomètre, faites à Paris pendant l'année 1735, comparées avec celles qui ont été faites sous la ligne, à l'isle de France, à Alger et quelques unes de nos isles de l'Amérique[M]. Paris: Memoires de l'Académie des Sciences de Paris, 1735 : 1 ~300.
(0)
|
| 31 |
Aono Y, Omoto Y. Estimation of blooming date for Prunus yedoensis using DTS combined with Chil 1-unit accumulations[J].
Journal of Agricultral Meteorology,1990, 45 (4) : 243~249.
(0)
|
| 32 |
Murray M B, Cannell M G R, Smith R I. Date of budburst of fifteen tree species in Britain following climatic warming[J].
Journal of Applied Ecology,1989, 26 (2) : 693~700.
(0)
|
| 33 |
Chuine I. A unified model for budburst of trees[J].
Journal of Theoretical Biology,2000, 207 (3) : 337~347.
(0)
|
| 34 |
王焕炯.
基于物候模型的中国木本植物春季物候变化研究[M]. 北京: 中国科学院大学硕士学位论文, 2012 : 1 ~61.
Wang Huanjiong. Shifts of Spring Phenology of Woody Plants in China Based on Phenological Models[M]. Beijing: The Dissertation for the Masteral Degree of University of Chinese Academy of Sciences, 2012 : 1 ~61. ( 0)
|
| 35 |
Možn Ay'3 M, Brázdil R, Dobrovoln Ay'3 P, et al. Cereal harvest dates in the Czech Republic between 1501 and 2008 as a proxy for March-June temperature reconstruction[J].
Climatic Change,2012, 110 (3-4) : 801~821.
(0)
|
| 36 |
郑景云, 葛全胜, 郝志新. 历史文献中气象记录与气候变化定量重建方法[J].
第四纪研究,2014, 34 (6) : 1186~1196.
Zheng Jingyun, Ge Quansheng, Hao Zhixin. Paleocliamtology proxy recoirded in historical documents and method for reconstruction climate change[J]. Quaternary Sciences,2014, 34 (6) : 1186~1196. ( 0)
|
| 37 |
Linkosalo T, Hkkinen R, Terhivuo J, et al. The time series of flowering and leaf bud burst of boreal trees(1846-2005)support the direct temperature observations of climatic warming[J].
Agricultural and Forest Meteorology,2009, 149 (3) : 453~461.
(0)
|
| 38 |
Aono Y, Moriya C. A generalized model to estimate flowering for cherry tree(Prunus yedoensis)considering both processes of endodormancy completion and development[J].
Journal of Agricultural Meteorology,2003, 59 (2) : 165~177.
(0)
|
| 39 |
Demarée G R, Chuine I.
A concise history of the phenological observations at the Royal Meteorological Institute of Belgium. In:Dalezios Nicolas R, Tzortzios Stergios eds. HAICTA 2006 Conference[M]. 2006 : 20 ~23.
(0)
|
| 40 |
Meier N, Rutishauser T, Pfister C, et al. Grape harvest dates as a proxy for Swiss April to August temperature reconstructions back to AD 1480[J].
Geophysical Research Letters,2007, 34 (20) : 1~6.
(0)
|
| 41 |
Cortázar-Atauri I G de, Daux V, Garnier E, et al. Climate reconstructions from grape harvest dates:Methodology and uncertainties[J].
The Holocene,2010, 20 (4) : 599~608.
(0)
|
| 42 |
Fu Y H, Piao S L, Zhao H F, et al. Unexpected role of winter precipitation in determining heat requirement for spring vegetation green-up at northern middle and high latitudes[J].
Global Change Biology,2014, 20 (12) : 3743~3755.
(0)
|
| 43 |
Gómez-Mendoza L, Galicia L, Cuevas-Fernández M L, et al. Assessing onset and length of greening period in six vegetation types in Oaxaca, Mexico, using NDVI-precipitation relationships[J].
International Journal of Biometeorology,2008, 52 (6) : 511~20.
(0)
|
| 44 |
Bradley A V, Gerard F F, Barbier N, et al. Relationships between phenology, radiation and precipitation in the Amazon region[J].
Global Change Biology,2011, 17 (6) : 2245~2260.
(0)
|
| 45 |
Yang Y G, Zhao C Y, Han M, et al. Temporal patterns of shrub vegetation and variation with precipitation in Gurbantunggut Desert, Central Asia[J].
Advances in Meteorology,2015 : 1~11.
doi: 10.1155/2015/157245 (0)
|
| 46 |
Zhu L, Meng J. Determining the relative importance of climatic drivers on spring phenology in grassland ecosystems of semi-arid areas[J].
International Journal of Biometeorology,2015, 59 (2) : 237~248.
(0)
|
| 47 |
Schaber J, Badeck F W. Physiology-based phenology models for forest tree species in Germany[J].
International Journal of Biometeorology,2003, 47 (4) : 193~201.
(0)
|
| 48 |
Blümel K, Chmielewski F M. Shortcomings of classical phenological forcing models and a way to overcome them[J].
Agricultural and Forest Meteorology,2012, 164 : 10~19.
(0)
|
| 49 |
Chen X Q, An S, Inouye D W, et al. Temperature and snowfall trigger alpine vegetation green-up on the world's roof[J].
Global Change Biology,2015, 21 (10) : 3635~3646.
(0)
|
| 50 |
Fu Y H, Zhao H F, Piao S L, et al. Declining global warming effects on the phenology of spring leaf unfolding[J].
Nature,2015, 526 (7571) : 104~107.
(0)
|
| 51 |
刘亚辰, 王焕炯, 戴君虎, 等. 物候学方法在历史气候变化重建中的应用[J].
地理研究,2014, 33 (4) : 1~11.
Liu Yachen, Wang Huanjiong, Dai Junhu, et al. The application of phenological methods for reconstructing past climate change[J]. Geographical Research,2014, 33 (4) : 1~11. ( 0)
|
| 52 |
Krinner G, Viovy N, Noblet-Ducoudré N de, et al. A dynamic global vegetation model for studies of the coupled atmosphere-biosphere system[J].
Global Biogeochemical Cycles,2005, 19 (1) : 1~33.
(0)
|
| 53 |
Sitch S, Smith B, Prentice I C, et al. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model[J].
Global Change Biology,2003, 9 (2) : 161~185.
(0)
|
| 54 |
Arora V K, Boer G J. A parameterization of leaf phenology for the terrestrial ecosystem component of climate models[J].
Global Change Biology,2005, 11 (1) : 39~59.
(0)
|
| 55 |
Delpierre N, Dufrêne E, Soudani K, et al. Modelling interannual and spatial variability of leaf senescence for three deciduous tree species in France[J].
Agricultural and Forest Meteorology,2009, 149 (6) : 938~948.
(0)
|
| 56 |
Sparks T H, Menzel A. Observed changes in seasons:An overview[J].
International Journal of Climatology,2002, 22 (14) : 1715~1725.
(0)
|
②. University of Chinese Academy of Sciences, Beijing 100049;
③. School of Geography, Beijing Normal University, Beijing 100875)
Abstract
Climate of the past is an important subject in the study of global change. Reconstruction of climate change by phenological records in history provides better understanding for the climate evolution stages in the past. In this study, we systematically summarized the plant phenological models used for climate reconstruction, including statistical models and process models. Statistical models establish regression equations between phenophases and climate factors based on their statistical relationship, ignoring the physiological process of plants. Process models simulate the growth of plants through the connection between the daily growth rate of plants and environmental factors based on semi-mechanism, mainly consist of SW(Spring Warming)model, DTS(the number of days transformed to standard temperature) model and model for simulating grape harvest dates. Subsequently, we introduced the applications of the mentioned models in quantitative climate reconstruction at various regions and discussed the accuracy and uncertainties of different models. At last, we put forward some key issues in reconstruction of past climate and suggest place importance on the reconstruction of other climate factors besides temperature. In the future, it is crucial to strengthen observations and studies on autumn phenology as well as to make thorough study of physiological process of plants in autumn, so that it is easier to learn the phenological process in autumn. Under such circumstances, there will be more choices in application of phenological models in the reconstruction of past climate, and we can make better use of the material in autumn for past climate reconstructions.